Guest column by Theo Heijnsdijk
Euphorbia clandestina was described in 1804 by Nicolas Joseph von Jacquin (1727 -1817) who was born and raised in the Dutch university town of Leiden. After studying medicine, he moved to Paris, and from there he ended up in Vienna. Apparently, natives of Leiden played an important role in Vienna at that time. The personal physician of Empress (of the Holy Roman Empire) Maria Theresa of Austria, Gerard van Zwieten, was a pupil of the famous Leiden physician Boerhaave. In 1753, on the recommendation of Van Zwieten, the gardener Adriaan Steckhoven, also from Leiden, was appointed director of the gardens of Schönbrunn Palace, also in Vienna. By order of Emperor Francis 1 Stephan, the husband of Maria Theresa, Von Jacquin was sent to Central America and the Caribbean to collect plants, animals, and minerals. The plant collection of the gardens was later described by Von Jacquin in the book “Plantarum rariorum horti caesarei Schoenbrunnensis descriptiones et icones” which appeared in four volumes between 1796 and 1804. Von Jacquin had long since left Schönbrunn. In 1762 he was appointed professor of minerals and mining at the Mining Academy in Schemnitz (now known as Banská Štiavnica). In 1768 he was appointed professor of botany and chemistry and became director of the botanical garden of the University of Vienna.
Outward appearance
In the aforementioned four-part series of books, 1270 plants are depicted and described, unfortunately in Latin, which is difficult to understand for most modern readers.
In volume 4 (1804) we find the description with colour image of E. clandestina.
 Fig. 1: The color plate accompanying Jacquin’s 1804 description.
Fig. 1: The color plate accompanying Jacquin’s 1804 description.
The slender stems are 2.5 to 4 cm in diameter and usually up to 60 cm high, but up to 1.2 m has also been reported. The leaves are up to 4 cm long and 2 to 4 mm wide. Older leaves drop off so that only a crown of leaves at the top remains. This gives a good view of the stem, which is covered in spirals of knobbly tubercles with a scar from the dropped-off leaf on the most outward part. See Figure 2.
 Fig. 2
Fig. 2
This appearance has given the species the name ‘volstruisnek’ (ostrich neck) in South Africa.
Flowering and fruiting
The species name clandestina (hidden, inconspicuous) refers to the inflorescence of the species. The greenish inflorescences (cyathia) that appear in spring are situated between the leaves at the top. As shown in figure 3 this produces a beautiful image.

Fig. 3: top view of a flowering plant.
In figure 4 we see a close-up of an individual cyathium in an early stage of flowering. The cyathophylls (bracts) are yellowish-green. They enclose a circle formed by 5 nectar glands. These in turn enclose the flowers proper. In the middle is the female flower (pistil) with 3 incised stigma lobes. On the left, we see a male flower (with the double anthers) that is just about to open and on the right male flowers that have already come out and with their tufts of stamens are somewhat reminiscent of a shaving brush. The species is self-fertile and because male and female flowers are often ripe simultaneously even in one cyathium, pollination is almost guaranteed.
 Fig. 4
Fig. 4
This results in fruits that are initially green and nearly spherical (fig. 5),
 Fig. 5 later turning into an indeterminate brownish shade (fig. 6);
Fig. 5 later turning into an indeterminate brownish shade (fig. 6); Fig. 6: An almost ripe fruit.
Fig. 6: An almost ripe fruit.
it can be seen that the remains of the stamens are still present). Once that stage has been reached, it is high time to take measures to prevent the seeds from being hurled away so that you are later confronted with “clandestine inhabitants” in the pots of other plants.
The natural habitat
E. clandestina is quite common in the Western Cape. There, the plants usually grow at altitudes between 60 and 450 meters, on hills among rocks and various kinds of thickets. In Figure 7, a photo by Coby Keizer, we see a specimen (estimated to be 1 meter tall) about 30 km north of the town of Riversdale.
 Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).
Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).
In Fig. 8, a photo by Bertus Spee taken at De Rust, we see what the sun in South Africa does to the plants: they are beautifully stocky and splendidly coloured.
 Fig. 8 Cultivation As far as cultivation is concerned, all sources say that the plant needs a light location and a lot of water and is certainly not very sensitive to cold. In the Western Cape, even light frosts seem to be tolerated. But under our climatic conditions, it is recommended not to allow the temperature to drop below about 5 °C. Sowing gives me varying degrees of success. On June 1, 2014, I sowed three rows of three freshly harvested seeds in a square pot. The three seeds in what I call the front row came up. See Figure 9.
Fig. 8 Cultivation As far as cultivation is concerned, all sources say that the plant needs a light location and a lot of water and is certainly not very sensitive to cold. In the Western Cape, even light frosts seem to be tolerated. But under our climatic conditions, it is recommended not to allow the temperature to drop below about 5 °C. Sowing gives me varying degrees of success. On June 1, 2014, I sowed three rows of three freshly harvested seeds in a square pot. The three seeds in what I call the front row came up. See Figure 9.  Fig: 9: Between the cotyledons of these 3 seedlings the first elongated leaves begin to develop. The other 6 didn’t. On June 10 I sowed 11, again freshly harvested seeds in a 5.5 cm pot. At the end of August, nothing had happened, and I put the pot under the tablet together with other pots with “unfinished business”. On September 20, more than 3 months later, I suddenly saw some greens under the table and it turned out that there were suddenly 7 seedlings present. On the same day, at the foot of the mother plants on the table, I also discovered some newly emerged seedlings. Weather conditions probably play a role here.
Fig: 9: Between the cotyledons of these 3 seedlings the first elongated leaves begin to develop. The other 6 didn’t. On June 10 I sowed 11, again freshly harvested seeds in a 5.5 cm pot. At the end of August, nothing had happened, and I put the pot under the tablet together with other pots with “unfinished business”. On September 20, more than 3 months later, I suddenly saw some greens under the table and it turned out that there were suddenly 7 seedlings present. On the same day, at the foot of the mother plants on the table, I also discovered some newly emerged seedlings. Weather conditions probably play a role here.
Similar species
E. clandestina is sometimes confused with E. clava which has stems of a similar form. That species has been known since the year 1700 but was only described around 1781, also by Von Jacquin. Fig. 10 shows the colour plate with the description.
 Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.
Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.
In the non-flowering state, both species can be distinguished because E. clava has slightly thicker trunks and produces branches from the base, whereas E. clandestina rarely branches, and if it does then not from the base.
But the most obvious is the difference in inflorescence. In E. clandestina, the inflorescences (cyathia) are sessile, that is, without a flower stalk. In E. clava, each cyathium always has a peduncle of 7.5 to 17.5 cm long. In Figure 11 we see a cyathium with a female flower. The flower stalks become woody and remain permanently attached to the plant like a kind of thorns.
 Fig. 11: Cyathium of Euphorbia clava with a female flower.
Fig. 11: Cyathium of Euphorbia clava with a female flower.
A species that is even more similar to E. clandestina is E. cylindrica. In this species, the stems are somewhat thicker (up to 5 cm) and the cyathia are raised on short stalks.
Literature:
Jacquin, N. J (1781 -1786): Icones plantarum rariorum Vol. I: 9, t 85.
Jacquin, N. J. (1804): Plantarum rariorum horti caesarei Schoenbrunnensis. Vol. IV: 43, t. 484 .
Laren, van, A,J. (1932). Succulents, Verkade’s factories N.V., Zaandam, NL.
Originally published in Succulenta 93 (6) 2014. Translated from Dutch by Frans Noltee.


 fig. 2
fig. 2


 Fig. 6. The seeds of A. papyracea are slightly prickly.
Fig. 6. The seeds of A. papyracea are slightly prickly. Fig. 7
Fig. 7



 Fig. 3
Fig. 3
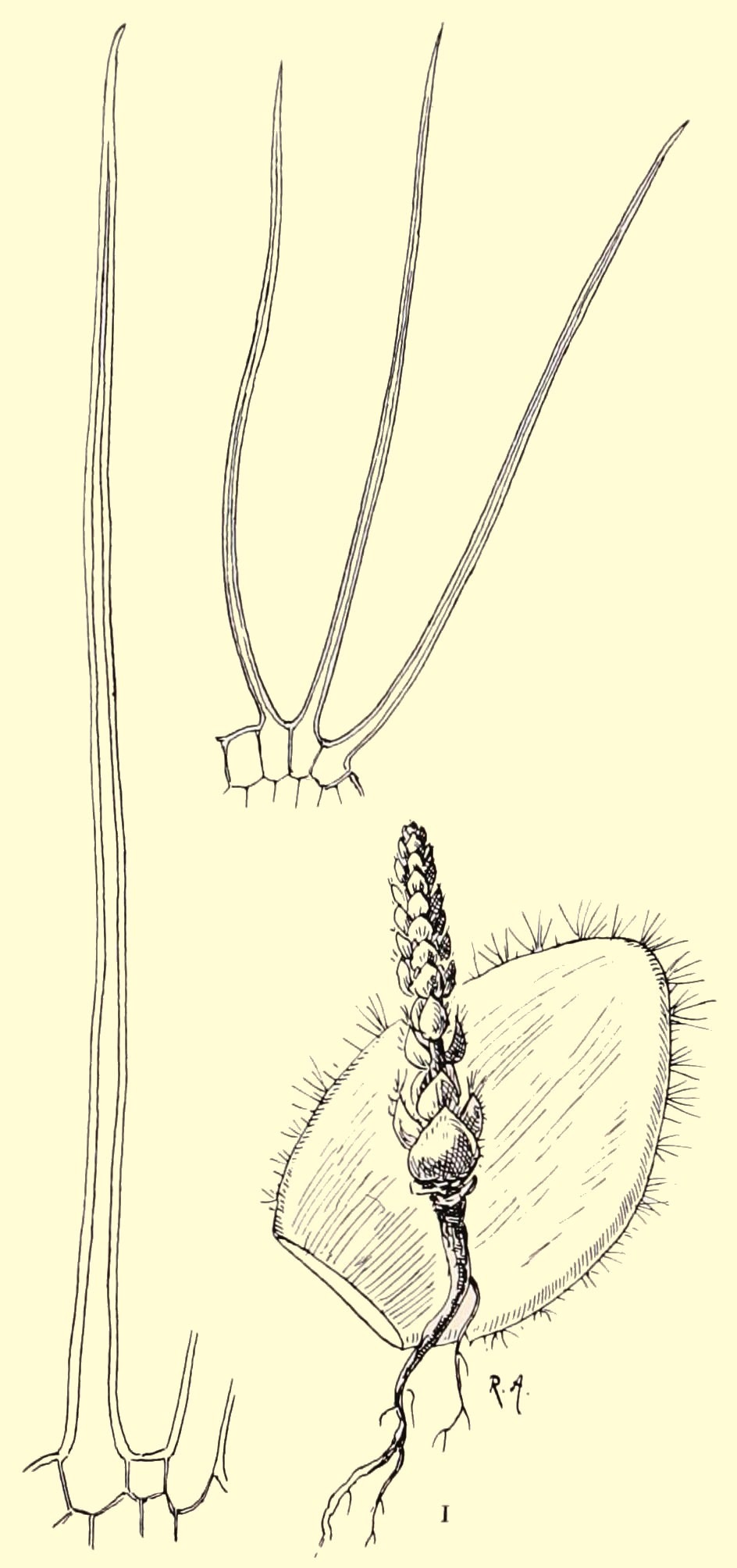
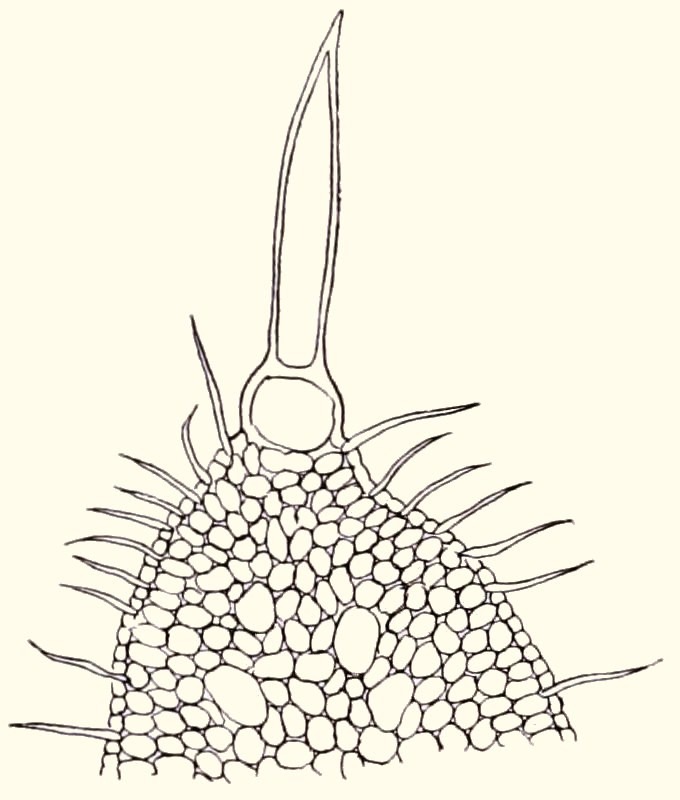





































 Fig 1
Fig 1
 Fig 4 C. columnaris ssp. prolifera in the Knersvlakte (photo by Sofia Etzold, with thanks to “Photo Guide to Plants of Southern Africa”). In Afrikaans, this variety is called ‘sentkannetjie’. ‘Sent’ in this expression is derived from the English ‘scent’ and kannetjie stands for bottle. This does justice to both the plant’s shape and the flowers’ sweet smell.
Fig 4 C. columnaris ssp. prolifera in the Knersvlakte (photo by Sofia Etzold, with thanks to “Photo Guide to Plants of Southern Africa”). In Afrikaans, this variety is called ‘sentkannetjie’. ‘Sent’ in this expression is derived from the English ‘scent’ and kannetjie stands for bottle. This does justice to both the plant’s shape and the flowers’ sweet smell.


















 Fig. 2: Crassula deceptor in bloom
Fig. 2: Crassula deceptor in bloom Fig 3.
Fig 3.

