Euphorbia clandestina was described in 1804 by Nicolas Joseph von Jacquin (1727 -1817) who was born and raised in the Dutch university town of Leiden. After studying medicine, he moved to Paris and from there he ended up in Vienna. Apparently, natives of Leiden played an important role in Vienna at that time. The personal physician of Empress (of the Holy Roman Empire) Maria Theresa of Austria, Gerard van Zwieten, was a pupil of the famous Leiden physician Boerhaave. In 1753, on the recommendation of Van Zwieten, the gardener Adriaan Steckhoven, also from Leiden, was appointed director of the gardens of Schönbrunn Palace, also in Vienna. By order of Emperor Francis 1 Stephan, the husband of Maria Theresa, Von Jacquin was sent to Central America and the Caribbean to collect plants, animals and minerals. The plant collection of the gardens was later described by Von Jacquin in the book “Plantarum rariorum horti caesarei Schoenbrunnensis descriptiones et icones” which appeared in four volumes between 1796 and 1804. Von Jacquin had long since left Schönbrunn. In 1762 he was appointed professor of minerals and mining at the Mining Academy in Schemnitz (now known as Banská Štiavnica). In 1768 he was appointed professor of botany and chemistry and became director of the botanical garden of the University of Vienna.
Outward appearance In the aforementioned four-part series of books, 1270 plants are depicted and described, unfortunately in Latin, which is difficult to understand for most modern readers.
In volume 4 (1804) we find the description with colour image of E. clandestina.
Fig. 1: The color plate accompanying Jacquin’s 1804 description.
The slender stems are 2.5 to 4 cm in diameter and usually up to 60 cm high, but up to 1.2 m has also been reported. The leaves are up to 4 cm long and 2 to 4 mm wide. Older leaves drop off, so that only a crown of leaves at the top remains. This then gives a good view of the stem, which is covered in spirals of knobbly tubercles with a scar from the dropped off leaf on the most outward part. See figure 2.
Fig. 2
This appearance has given the species the name ‘volstruisnek’ (ostrich neck) in South Africa.
Flowering and fruiting The species name clandestina (hidden, inconspicuous) refers to the inflorescence of the species. The greenish inflorescences (cyathia) that appear in spring are situated between the leaves at the top. As shown in figure 3 this produces a beautiful image.
Fig. 3: top view of a flowering plant.
In figure 4 we see a close-up of an individual cyathium in an early stage of flowering. The cyathophylls (bracts) are yellowish green. They enclose a circle formed by 5 nectar glands. These in turn enclose the flowers proper. In the middle is the female flower (pistil) with 3 incised stigma lobes. On the left we see a male flower (with the double anthers) that is just about to open and on the right male flowers that have already come out and with their tufts of stamens are somewhat reminiscent of a shaving brush. The species is self-fertile and because male and female flowers are often ripe simultaneously even in one cyathium, pollination is almost guaranteed.
Fig. 4
This results in fruits that are initially green and nearly spherical (fig. 5), Fig. 5
later turning into an indeterminate brownish shade (fig. 6;
Fig. 6: An almost ripe fruit.
it can be seen that the remains of the stamens are still present). Once that stage has been reached, it is high time to take measures to prevent the seeds from being hurled away so that you are later confronted with “clandestine inhabitants” in the pots of other plants.
The natural habitat
E. clandestina is quite common in the Western Cape. There the plant usually grows at altitudes between 60 and 450 meters, on hills among rocks and various kinds of thickets. In figure 7, a photo by Coby Keizer, we see a specimen (estimated to be 1 meter tall) about 30 km north of the town of Riversdale.
Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).IMG_4806 Coby Keizer.
In Fig. 8, a photo by Bertus Spee taken at De Rust, we see what the sun in South Africa does to the plants: they are beautifully stocky and splendidly coloured.
Fig. 8.
Cultivation
As far as cultivation is concerned, all sources say that the plant needs a light location and a lot of water and is certainly not very sensitive to cold. In the Western Cape, even light frosts seem to be tolerated. But under our climatic conditions, it is recommended not to allow the temperature to drop below about 5 °C.
Sowing gives me varying degrees of success. On June 1, 2014, I sowed 3 rows of 3 freshly harvested seeds in a square pot. The 3 seeds in what I call the front row came up. See figure 9.
Fig: 9: Between the cotyledons of these 3 seedlings the first elongated leaves begin to develop.
The other 6 didn’t. On June 10 I sowed 11, again freshly harvested seeds in a 5.5 cm pot. At the end of August, nothing had happened, and I put the pot under the tablet together with other pots with “unfinished business”. On September 20, more than 3 months later, I suddenly saw some greens under the tablet and it turned out that there were suddenly 7 seedlings present. On the same day, at the foot of the mother plants on the table, I also discovered some newly emerged seedlings. Weather conditions probably play a role here.
Similar species E. clandestina is sometimes confused with E. clava which has stems of a similar form. That species has been known since the year 1700 but was only described around 1781, also by Von Jacquin. Fig. 10 shows the colour plate with the description.
Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.
In the non-flowering state, both species can be distinguished because E. clava has slightly thicker trunks and produces branches from the base, whereas E. clandestina rarely branches and if it does then not from the base.
But most obvious is the difference in inflorescence. In E. clandestina, the inflorescences (cyathia) are sessile, that is, without a flower stalk. In E. clava, each cyathium always has a peduncle of 7.5 to 17.5 cm long. In figure 11 we see a cyathium with a female flower. The flower stalks become woody and remain permanently attached to the plant like a kind of thorns.
Fig. 11: Cyathium of Euphorbia clava with female flower.
A species that is even more similar to E. clandestina is E. cylindrica. In this species, the stems are somewhat thicker (up to 5 cm) and the cyathia are raised on short stalks.
Literature: Jacquin, N. J (1781 -1786): Icones plantarum rariorum Vol. I: 9, t 85.
Jacquin, N. J. (1804): Plantarum rariorum horti caesarei Schoenbrunnensis. Vol. IV: 43, t. 484 .
Laren, van, A,J. (1932). Succulents, Verkade’s factories N.V., Zaandam, NL.
Originally published in Succulenta 93 (6) 2014. Translated from Dutch by Frans Noltee.
Taxonomy
Avonia used to be a subgenus of Anacampseros, a relatively small genus in the family of purslanes (Portulacaceae).
The species within the genus Anacampseros consist of small to very small shrublets with a more or less thick, sometimes branched stem. The fleshy leaves form a flat rosette or are attached to the stem in a spiral. A typical characteristic are the stipules (lateral outgrowths at the base of the leafstalks). Often these have been reduced to hairs, or they have grown into parchment-like white scales which in some species can completely cover the stem including the real leaves. In 1994 Gordon Rowley concluded that these species with scale-shaped stipules deserve the status of a separate genus and thus the genus Avonia came into existence. In it he placed 9 species. There are now 12. The name is probably derived from the Latin ‘Avus’ which means grandfather. It would then refer to the white scales that give the plants an ‘old’ appearance. Rowley placed a few other Anacampseros-like plants in the genus Grahamia. For the genus Anacampseros itself, according to his classification, 15 species remain.
Together, the genera Anacampseros, Avonia and Grahamia form the branch (tribe) Anacampseroteae within the Portulacaceae.
Incidentally, it is doubtful whether the genus Avonia will hold. It is rather unusual in this day and age to base a genus on a similarity in its external appearance. As a rule, characteristics of the flower and seed and even more so the similarities and differences in DNA play the decisive role.
Flower and fruit
What all representatives of the Anacampseroteae have in common is the construction of flower and fruit. The flowers, which in all species are only open for a few hours in the late afternoon, always have only 2 sepals, which first completely enclose the flower and often curl up after flowering but remain joined at the tip so that they stay on top of the developing fruit like a cap. This can be seen very well in Avonia quinaria ssp. alstonii (fig.1). At a certain moment the cap falls off and the seeds are released.
Fig. 1
Furthermore, there are usually 5 (sometimes 4) petals , which are white, pink or carmine. The stamens are white with yellow anthers and very variable in number: from 5 in, for example, Anacampseros comptonii to as much as 80 in A. quinaria (fig. 2). The white pistil has 3 lobes. The fruit consists of a kind of veined basket that often has a narrow opening at the top, so that the seeds come out in dribs and drabs as if with a salt-sprinkler as the flower stem bends to the ground and is moved up and down by the wind.
Fig 2. Avonia quinaria ssp. alstonii has the largest number of stamens of all Anacampseroteae (up to 80).
A. papyracea was chosen as the type species of the genus because of its most pronounced characteristics. The name papyracea (=paper-like) is particularly appropriate because a stem really looks like a little wad of wafer-thin pieces of paper. The name was given by the German botanist and director of the botanical garden in Königsberg, Ernst Heinrich Friedrich Meyer (1791-1858). The description in accordance with the taxonomic rules followed a bit later and is by the Austrian botanist Eduard Fenzl. It appeared in 1840 in a large article about the Molluginaceae (which at that time still included the Portulacaceae) in ‘Annalen des Wiener Museums der Naturgeschichte’. Hence the designation ‘E. Meyer ex Fenzl as found in botanical literature.
The white scales undoubtedly protect the green leaves underneath them against the bright sunlight. Rowley observed that under humid conditions the scales deflect somewhat from the stem so that there can be a better exchange of gases (oxygen, water vapor) with the environment. Perhaps water can also be absorbed in liquid form.
Like most Avonias, A. papyracea hails from South Africa (Western and Northern Cape). It is fairly common and grows mainly in plains with white quartz stones. But the plant also grows on dark coloured soil, where it is of course much more obvious. Figure 3, a photograph by Coby Keizer, shows a cluster in Goegap Nature Reserve east of Springbok.
Fig. 3
The former director of the Royal Botanic Gardens at Kew, Sir William T. Thiselton–Dyer, pointed out that the white stems bear a certain resemblance to bird droppings. It is not for nothing that the plant is also called ‘gansmis’ (goose dung) in South Africa. Probably the geese have a different menu there than in Europe, because here the droppings of geese are just as green as the grass they eat. Thiselton–Dyer suggested that this is a mimicry plant, which in this way tries to prevent it from being seen as a tasty snack. If that is the case, then it does not seem to be a successful imitation, because another name for the plant is ‘haasieskos’ (Hare food). The same name is also used for A. ustulata and A. filamentosa. Thiselton-Dyer made his remark in 1906 in an article in the British ‘Annals of Botany’. At that time, the flower was still completely unknown. In the same article he mentions that his friend Nicholas Edward Brown, also from Kew, had been growing the plant for many months, but despite daily observation had never seen a flower on it. Yet, suddenly a fruit appeared from between the parchment-like leaves. He concluded that the flower had to be very small and hidden under the scales had fertilized itself. A cleistogamous species, in other words. Later authors also maintain that the flowers do not open. But that’s not true. Sometimes the plant does bloom and the flowers are not small nor do they remain hidden under the scales (Fig. 4).
Fig. 4. The flower of A. papyracea and a seed pod.
Thiselton-Dyer also reported in the article mentioned above that the stalk of the ripening fruit continues to grow so that the seeds are scattered at some distance from the mother plant. This can also be clearly seen in Fig. 4. However, this is not always the case. In Fig. 5 it can be seen that the 2 fruits hardly appear from between the scales.
Fig. 5. A full seed pod of A. papyracea and an already emptied one. Both only just visible between the scales.
In addition to the usual species which is nowadays referred to as A. papyracea ssp. papyracea, there is also a subspecies: A. papyracea ssp. namaensis. This occurs in the northwestern part of South Africa and in the south of Namibia and is distinguished by the fact that the scales are not smooth-edged but serrated (saw-edged) or toothed and by the seeds that are lumpy. In ssp. papyracea they are more prickly (Fig. 6).
Fig. 6. The seeds of A. papyracea are slightly prickly.
Seed collection is extremely simple. Holding an empty tube or something similar under a ripe fruit and a tap against the fruit are enough. Immediate sowing gives excellent results. At a temperature of 25 to 30 °C, the seedlings emerge en masse after a week. Only a short time later, the seedlings proceed to develop the papery scales (Fig. 7).
Fig. 7
Fig. 8. After a year, they are miniature versions of the mature plants.
Cultivation
I grow the plants in standard cactus soil and I also treat them like cacti. This means that they receive water regularly in summer and that I keep them dry in winter. I try to keep the temperature in my greenhouse around 8 °C in winter, but it sometimes drops to just above freezing point. According to the literature, even temperatures of 5 degrees below zero are survived.
Apparently, A .papyracea, like A. ustulata, was also used in South Africa as a kind of yeast for baking bread and making beer. To this end, roots and stems are dried and ground. It is suspected that the plant also contains psychoactive substances, just as, by the way, A. quinaria. The brew must have been a kind of ‘spacebeer’.
Literature
Browne, P. (1756). The Civil and Natural History of Jamaica. In Three Parts [3]: 234
Fenzl, E. (1840). Monographie der Mollugeneen, Annalen des Wiener Museums der Naturgeschichte, [2]: 295
Marloth, R (1917). Dictionary of The Common Names of Plants, The Specialty Press of South Africa, Ltd. Cape Town
Rowley, G. (1994). Anacampseros and allied genera- A reassessment, Bradleya 12: 105-112
Sims, J. (1811) Anacampseros filamentosa, Curtis’s Botanical Magazine 33: plate 1367
Thiselton-Dyer, W.T.(1906). Morphological Notes, Annals of Botany 20 (2) blz 123-127
Originally published in Succulenta 93: 2014. Translated from Dutch by FN.
C. barbata is among the 28 new South African Crassula species described by Carl Thunberg in 1778. In nature it occurs on rocky slopes in quite a large area including, among others, the Great Karoo and the Little Karoo. Characteristic for the species are the up to 5 millimeter long hairs on the leaf margins which give the impression of entangled eyelashes (fig. 1) or a beard – as the the botanical name implies (barbata = with a beard).
fig. 1 At the leaf tips, the hairs are often gathered in tufts (fig. 2). Fig. 2 Apart from these long hairs, the leaves are completely hairless. At first glance, especially in the resting period, these spheres look like ball-shaped cacti such as Mammillaria plumosa. See fig. 3, a photo made by Coby Keizer in November 2008 at the farm Perdekraal, circa 40 km west-northwest of Matjiesfontein. There are a lot of very interesting succulent plants there. Frans Noltee wrote two articles about it in Succulenta in 2008. The small other plants (upright fingers) in fig. 3 are probably Cephalophyllum curtophyllum.fig. 3
In the guest column on CRASSULA PYRAMIDALIS * I mentioned the experiments that Rudolph Marloth carried out early last century to asses water absorption by Crassulas through the leaves and hairs. A C. barbata gained 5.2% in weight in one night by absorbing dew. Marloth also looked at what happens when you apply a small drop of water to the end of a hair on a wilting leaf. Before long, the hair swells and stands upright and gradually the leaf regains its turgor. In Crassula tomentosa (fig. 4), with its row of thick hairs, this works even better. Marloth took a leaf from it and dipped the hairy edge in the water. After 12 hours, the weight had increased from 14.45 to 16.05 gram, an increase of no less than 11%.
fig.4 Marloth’s article was illustrated with several beautiful drawings in which you can also see that the hairs have developed from epidermis cells (fig. 5 and fig. 6).
fig. 5fig. 6
C. barbata is a slow grower. It takes a few years until the heart of the rosette opens and the beginning of the flower stem becomes visible (in the Netherlands usually at the end of November). At that stage, that flower stem looks like a C. pyramidalis plantlet (fig. 7).
fig. 7 The stem grows rapidly (fig. 8 and fig. 9) and after a few weeks the maximum height (up to 30 cm) is reached.
fig. 8
fig. 9 At the bottom there are still leaves with lashes. The flowers are arranged in groups of about 7 and several of those groups stand in a crown around the stem. The petals are usually beautifully white and this contrasts nicely with the yellow anthers (fig. 10).
fig. 10
Pink shades also occur (fig.11).
fig. 11 The flowers emit a pleasant sweet smell.
There is only one known variant of C. barbata: ssp. broomii, also known as an independent species: C. broomii. It grows in the vicinity of Victoria West (North Cape) in the mountains and is distinguished from the type plant mainly because the hairs are considerably shorter, up to only 1 mm. For plantlovers this makes it not so interesting, because it is precisely the hairs that make the plant so attractive. We therefore hardly ever encounter this variety in collections.
There are also hybrids of C. barbata, but they are not common in culture. In an article in Succulenta (1980), B.K. Boom mentions a cross with C. orbicularis cv. ʽRosula’ and also shows a photo. Rowley mentions a cv. “Roger Jones” as being “C. barbata x ? orbicularis”, but does not give an image.
Gordon Rowley, in his book ‘Crassula’, considers C. barbata a joy to see in its natural habitat but usually a disappointment in cultivation. Instead of remaining compact, the plants grow into elongated loose rosettes and after flowering the plant dies. But he also made this kind of statement about C. columnaris and C. pyramidalis. My experience is that it is not so bad as long as you are economical with water. Watch out for total dehydration in summer and give the plant the lightest spot you can find in winter.
Also, the plant does not always die after flowering. For example, I had a plant that bloomed in the winter 2011-2012. As the rosette died off, 5 new shoots formed at the base of this plant. At the moment, at the beginning of December 2013, a flowering stem rises above the rosette in all 5 0f them. So you can enjoy such a plant for years.
Literature: Boom, B.K. (1980). De Crassulas van onze collecties 6 and 7, Succulenta 59 (1): 120. Laren, van, A,J. (1932). Vetplanten, Verkade fabrieken N.V., Zaandam. Marloth, R. (1908). Das Kapland, Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer “VaIdivia” 1898-1899, bd.2, t.3. Noltee, F. (2008). Succulenten van de Karoo, een bezoek aan Perdekraal, Succulenta 87 (4): 170 and Succulenta 87 (5): 215. Rowley, G. (2003). Crassula, Cactus & Co. Thunberg C.P. (1778). Nova acta physico-medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum 6: 328-341.
Originally published in Succulenta 93 (4) 2014. Translated from Dutch by Frans Noltee.
The year 1778 is an important one for Crassula lovers. In that year, an article appeared in the Swedish periodical with the wonderful name ‘Nova Acta Physico-Medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosum’ in which physician-botanist Carl Peter Thunberg (also from Sweden) briefly described no less than 28 new South African species of this genus. Most of these have become well-known among fanciers. Just think of Crassula alpestris, C. barbata, C. capitella, C. columnaris, C. corallina, C. hemisphaerica, C. tecta, C. tomentosa, and the subject of this article: C. pyramidalis.
At the back of Thunberg’s book is a plate depicting 2 of the new species, C. alpestris and C. pyramidalis (see fig. 1).
fig. 1
In later years, of course, many more images of C. pyramidalis appeared. Particularly beautiful is the coloured plate published in 1899 in Curtis’s Botanical Magazine (fig. 2). Here we see a thicker shape than Thunberg’s plant has.
fig. 2
C. pyramidalis is closely related to C. columnaris (discussed earlier in this series) and to C. barklyi. All three of them are highly succulent species whose leaves are so tightly packed together that the stem is totally invisible. In C. pyramidalis, the leaves are grouped very regularly. At the top of a stem, this results in a particularly beautiful view (fig. 3).
fig. 3
In nature, the plants can take on a beautiful purple-red color (See figure 4). Coby Keizer took this picture in November 2008 about 60 km north of Matjiesfontein (Western Cape) along the road to Sutherland.
fig. 4
Not visible in photos are the hairs in the leaf axils. They allow the plant to absorb water. In the Dutch picture album of the Verkade Company this is described by the author, A. J. van Laren, as follows: ‟Remarkable in some of these Crassulas is the ability to absorb water by means of hairs on the leaves, such as the marginal hairs of C. barbata; in C. columnaris and C. pyramidalis there are such water-absorbing hairs at the base of the leaves, thus at the trunk. The effectiveness of these hairs is rather significant! To give an example, a leaf of C. barbata can absorb more water in one night, as a result of dew, than it loses in a week by transpiration. The placement of the leaves in C. pyramidalis is such that water flowing down the plant penetrates into the spaces between the leaves, from where it is channeled to the base and absorbed by the hairs present there. When drought sets in, the leaf margins are so close together that the water-absorbing hairs at the base of the leaves are completely closed off from the outside air and therefore cannot lose water.
I suspect that van Laren derived the above information from the book which the German chemist/pharmacist/botanist Rudolph Marloth (1855 -1931) wrote under the title ‘Das Kapland, insonderheit das Reich der Kapflora, das Waldgebiet und die Karroo’. This book with over 550 pages in 1908 as part 3, volume 2 of a whole series under the title ‘Wissenschaftliche Ergebnisse der Deutschen Tiefsee-expedition auf dem Dampfer “VaIdivia” 1898-1899’. The Valdivia was a steamer commissioned by the German government to undertake a deep-sea expedition in the Atlantic and Indian Oceans from August 1898 to May 1899. The report under the title mentioned above included 24 volumes, the first of which appeared in 1902 and the last in 1940. Marloth, who lived and worked in South Africa from the age of 28 until his death, did not participate in the expedition but became involved by the botanist Wilhelm Schimper who disembarked on the return trip to Cape Town to continue with botanical fieldwork. The concept of deep-sea was apparently understood very broadly. When Schimper died in 1901, Marloth was asked to complete the volume about South Africa.
In his book, Marloth reports on experiments with various Crassula species. He took 2 specimens of C. pyramidalis of which he wrapped the roots in tin foil so that they were closed off from the open air. He put the plants outside for a night. One covered with a piece of cardboard, the other one unprotected. The covered specimen lost a bit of weight (0.6%), whereas the unprotected specimen exposed to the dew increased in weight by 5.1%. Marloth concluded that the plant can absorb 5.7% of its body weight from dew. Of course, that does not prove at all that this is caused by those leaf axil hairs, but it is still a nice fact. According to Marloth, C. columnaris also has such hairs in the leaf axils. In his experiment, this species was found to experience 6.3% weight gain due to the dew. For C. barbata, the increase was 5.2%.
In addition to the position of the leaves, C. pyramidalis, C. columnaris and C. barklyi also correspond in the terminal, almost spherical inflorescence without visible flower stem (fig. 5) so that it seems as if a globule of flowers is crammed into the plant from above. The visual disadvantage is that the dried plug remains in place for a long time after flowering.
fig. 5
C. pyramidalis tends to allow all stem ends to bloom at once. A beautiful sight, but also the harbinger of a rapidly approaching death. This happened to me, for example, with the plant in fig. 6 that I bought at an open day at the Ubink nursery.
fig. 6 The photo was taken 3 months after the purchase. The stems were nicely thickened and coloured slightly red by the sun. At the end of February 2012 I saw that the formation of buds had started (fig. 7).
fig. 7
On April 1, the first flowers were open and on April 9, all stems were in bloom (fig. 8). The flowering lasts for many weeks, but the flowers gradually turn brown, after which the same thing happens to the stems themselves and then the plant is done for.
fig. 8
Thunberg found C. pyramidalis at the thermal springs on the Olifants River (Cape Province). Later, the species was found in many other places in the Cape Province (Little Karoo, Great Karoo, Namaqualand). The species is widespread in other words.. We can also come across the plant in literature under one of the following names: Crassula quadrangula, Tetraphyle pyramidalis, Tetraphyle quadrangula, Tetraphyle pyramidalis var. quadrangula, Purgosea pyramidalis, Crassula archeri and Crassula cylindrica.
In his 1778 article Thunberg wrote that C. pyramidalis resembles C. muscosa, but is distinguished by the thicker stems and the terminal inflorescence. C. muscosa is the valid name for the plant that probably most of us know as C. lycopodioides, the well-known shoelace plant. In this species, the inconspicuous and (to my mind) unpleasantly smelling flowers appear everywhere along the stems between the tiny leaves (fig. 9). Muscosa, by the way, means: moss-like, referring to the collection of thin stems. Lycopodioides means: resembling wolf’s claw (Lycopodium).
fig. 9
Over the years, a number of hybrids of C. pyramidalis have become available. Without a doubt the best known is ‘Buddha’s Temple’, a hybrid with C.perfoliata var. falcata. A photo of this plant in side view was already shown in the article in this series about the latter. In fig. 10 this plant is shown in top view.
fig. 10
We also know ‘Giant’s Watch Chain’ (C. muscosa x pyramidalis), ‘Starburst’ (C. ausensis x pyramidalis), and the multihybrides ‘Star Child’ (C. deceptor x ‘Starbust’) and ‘Tinkerbell’ (C. ausensis x ‘Springtime’) x pyramidalis) in which ‘Sprintime’ in its turn is a cross between C. perfoliata var. falcata with (probably) C. rupestris. A hybrid of no less than 4 species. It will probably make your head spin in the meantime.
A hybrid that also deserves a place here is C. ‘Extagnol’ which we also come across under the name C. ‘Estagnol’. This plant stands out because of the spiraling leaf position. In close-up (fig. 11) it reminds one somewhat of Aloe polyphylla .
fig.11
But remember that the diameter of this plant is only 10 mm. Its origin is not clear, but in the literature it is stated that it is a cross between C. pyramidalis and C. perfoliata var. perfoliata. I think the leaf shape is more indicative of C. alpestris, but that is only a random guess of course.
In cultivating C. pyramidalis and all these hybrids, we must bear in mind the highly succulent character of the plants. That is to say: a mineral soil, a sunny location, careful with water and little to no fertilizer.
Literature
Heijnsdijk, T. (2013). Crassula perfoliata var. falcata, Succulenta 92 (1): 3.
Heijnsdijk, T. (2013). Crassula barklyi, Succulenta 92 (4): 178.
Hooker, J. D. (1899). Curtis’s Botanical Magazine 125, tab 7665.
Laren, van, A,J. (1932). Succulents, Verkade’s factories N.V., Zaandam.
Marloth, R. (1908). Das Kapland, Wissenschaftliche Ergebnisse der Deutschen Tiefsee-expedition auf dem Dampfer “VaIdivia” 1898-1899, bd.2, t.3.
Thunberg, C.P. (1778). Nova acta physico-medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum, t6: 328-341
Images:
Fig. 1 The first published image of C. pyramidalis (Thunberg, 1778)
Fig. 2 A part of the image in Curtis’s botanical from 1899
Fig. 3 A young side shoot
Fig. 4 C. pyramidalis approx. 60 km north of Matjiesfontein along the road to Sutherland. (photo Coby Keizer)
Fig. 5 The almost spherical inflorescence of C. pyramidalis.
Fig. 6 C. pyramidalis in a 5 cm pot
Fig. 7 Button formation at the plant of fig. 5 C. pyramidalis
Fig. 8 The same plant: all shoots bloom at the same time
Fig. 9 Crassula muscosa (lycopodioides) in bloom
Fig. 10 Crassula ‘Buddha’s Temple’ (C. perfoliata var. falcata x pyramidalis)
Fig.11 Crassula ‘Extagnol’ (possibly a hybrid of C. pyramidalis and C. perfoliata var.perfoliata)
Originally published in Succulenta 93 (3) 2014. Translated from Dutch by Frans Noltee.
Succulent fanciers usually are not impressed by Haworthia flowers. On a wispy stalk there are a number of insignificant, mostly dirty white flowers. In H. truncata, the flowers have brownish longitudinal stripes. How different things become when you look at one of those flowers in close-up (fig. 1).
Fig. 1 Close-up of a flower of Haworthia truncata
The petals end in gracefully curly transparent slips and the ensemble is reminiscent of a majestic flying swan.
In nature the pollination of the tubular flowers is often done by bee species with extra-long mouthparts. Honey suckers (the African counterpart of the hummingbird) also play a role in pollination.
Fig. 2 The plate accompanying the description by Schönland in Transactions of the Royal Society of South Africa (1910).
Haworthia truncata was found in April 1909 by a certain Miss L. Britten on a farm 7 miles from the town of Oudtshoorn in the Little Karoo (Western Cape, South Africa). In 1910, the description by botanist Selmar Schönland appeared in the South African magazine ‘Transactions of the Royal Society of South Africa’. The accompanying plate shows the plant with detailed drawings of the various parts (see fig. 2).
Fig. 2 The plate accompanying the description by Schönland in Transactions of the Royal Society of South Africa (1910).
The formal description is followed by Schönland’s reflection on the morphology and anatomy of this special plant.
Karl von Poellnitz distinguished three forms of H. truncata in 1938: fa. normalis, fa. crassa with thicker and less flat leaves and fa. tenuis which remains much smaller. The latter was described as a variety by M.B. Bayer in 1976.
According to current taxonomic insights, these different names are unjustified and one name is sufficient for all the varieties and forms mentioned: H. truncata subsp. truncata. The closely related H. maughanii is seen as another subspecies: H. truncata subsp. maughanii.
Haworthia truncata is an outsider among the Haworthias. To begin with, the species differs from the standard Haworthia in that the leaves are not arranged in a rosette but are aligned (see fig. 3).
Fig. 3 Seedling of Haworthia truncata (Vanwykskraal) in cultivation.
In addition, the leaves, which are almost completely hidden in the ground, are flattened at the top (but with some differences in height) and all at about the same level so that it looks like they have been cut off with a blunt knife just above the ground (truncata means shortened). The cross-section is almost rectangular, so that the whole plant (sideways) is reminiscent of a fan or a multi-armed candlestick. The upright truncated leaves have earned the plant the name ‘Perdetande‘ (Horse Teeth) in South Africa. In Schönland’s drawing (fig. 2) the leaves do not have that typically rectangular shape, but are more similar to the leaves of H. maughanii.
To prevent the growing plant from protruding above the ground, there is another special faculty. The roots have transverse grooves that allow them to contract and thus bring down the plant body. Pretty much like an earthworm can contract.
Schönland explains that the plant, of which only the top of the leaves is visible, looks like a collection of pebbles and therefore does not stand out. In other words: it is a mimicry plant that in this way tries to protect itself from being eaten by animals. In order to receive enough light for assimilation, the leaf surface is somewhat transparent so that the light can penetrate to the assimilating tissue located on the inside of the leaf. This means that H. truncata can be counted among the so-called window plants, which we find mainly in the mesems. Think of Fenestraria, Frithia and several Lithops and Conophytum species. Schönland believes that this type of Southern African plant species that grow in full sun should in fact physiologically be considered to be shade plants.
In South Africa, they use different wording for all this.
In the question and answer section ‘Vra vir Ernst’ of a South African newspaper, I found the following description of H. truncata by Ernst van Jaarsveld:
‟In nature, the Horse Teeth are found in the Klein Karoo, especially in the Oudtshoorn district. They grow partly under bushes in stony conglomerate soil. Only the blunt leaf tips stick out of the ground, like horse teeth. They are difficult to spot and adapted to the arid environment. The leaves like those of beeskloutjies (little cow hoofs = Lithops spp.) are translucent green. In times of drought, the leaves shrink, and the dust covers the plants until they are almost invisible to humans.”
Haworthia truncata remained rare in European collections for quite some time after its discovery in 1909. In the Dutch monthly magazine ‘Succulenta’ it was first written about only in 1932 by the then widely known G.D. Duursma:
“Haworthia truncata Schönl. is a welcome addition of recent years, predestined to become so popular that it will soon be present in many collections.”
Cultivating the species is not particularly difficult. In South Africa, the plant grows and flowers in the rainy season. That’s in September and October. It makes sense to maintain this growing period in the northern hemisphere as well. This means that the plant should receive as much light as possible in these months and be watered regularly. In our climate, growth usually takes place in spring too. In the summer months, shade should be provided in the heat of the day and just enough water should be given to prevent the plant from drying out. The pot should not be too large, because then the soil may remain wet for too long after watering, resulting in rot. In literature it is reported several times that the roots periodically die and that the plant then quickly makes new roots in fresh soil. Like all plants of the Western Cape, the species is not sensitive to cold.
As a rough estimate, H. truncata grows at less than 10 localities in the vicinity of Oudtshoorn and west of it near Calitzdorp. Well-known locations are Dysselsdorp and De Rust, both east of Oudshoorn.
Fig. 4 Haworthia truncata near Dysseldorp (photo Coby Keizer).
Fig. 4 shows a group on a flat hilltop west of Dysselsdorp. The plants there occur generally in full sun but sometimes also under bushes. The soil is sandy and mixed with large and small stones.
Other succulents Coby has seen there are Cotyledon orbiculata, Aloe humilis and Aloe humilis hybrid (humilis x microstigma), Anacampseros arachnoides, Crassula capitella ssp.thyrsiflora, Crassula subacaulis, Duvalia species, Quaqua spec., Stapelia spec, Gasteria spec, various mesems and caudiciform plants. A true paradise for succulent fanciers.
Between Dysselsdorp and Kammanassie Dam (about 10 km to the south), a small variation occurs in which the leaf edges usually have small, pointed protrusions reminiscent of hairs. This form was described by German Ingo Breuer as var. minor (fig. 5, photo Robert Wellens). Later on, Breuer elevated this variety to a separate species: H. papillaris.
Fig. 5 Haworthia truncata var. minor VA 6718 (photo Robert Wellens).
At Van Wykskraal, about 5 km from Dysselsdorp, a natural hybrid with H. arachnoidea seems to grow.
The populations have suffered severely from the collecting frenzy of succulent lovers and traders. In addition, there is habitat destruction due to increasing urbanization of the area. On the Red List of endangered South African plants, the species has been given the status ‘vulnerable’.
In addition to the natural hybrids, there are countless artificial ones, often with variegated leaves or different folds of the leaf surface. Worth mentioning is the hybrid “Lime Green” (fig. 6, photo Robert Wellens), probably a cross with H. cuspidata, although H. cymbiformis is also considered to be a possible parent.
Fig. 6 Haworthia ‟Lime Green” (photo Robert Wellens).
Of H. maughanii (fig. 7-9) only one locality, south of Calitzdorp, is known. The area is less than 1 km2. Here too, a lot of damage has been done by succulent hunters and there is also damage from ostriches trampling the plants. All this has led to the status of ‘Critically Endangered’ for this species. The habitat of this species partly overlaps with that of the form of H. truncata which was described as fa. crassa and there are all kinds of natural hybrids in this area.
Fig. 7 A seedling of Haworthia maughanii (H. truncata ssp. maughanii) in cultivation.Fig. 8 Haw. truncata v. maughanii in habitat. Photo Frans Noltee
| Fig. 9 Haw. truncata v. maughanii in habitat. Photo Frans Noltee
Literature:
–Duursma, G.D. (1932) Haworthia truncata, Succulenta 14 (7): 169-172.
–Jaarsveld, E. van (2001). Vra vir Ernst, Lastige molkrieke en die biologiese bekamping van plae, Die Burger, Kultuurkroniek, http://152.111.1.87/argief/berigte/dieburger/2001/09/08/4/19.html
–Schönland, S. (1910). On some points in the morphology and biology of a new species of Haworthia, Transactions of the Royal Society of South Africa 1 (2): 391-394.
–Red list of South African Plants, http://redlist.sanbi.org/genus.php?genus=2215
First published in Succulenta 93, 2014-2. Translated from Dutch by Frans Noltee.
In 1897, professor Peter MacOwan, a British botanist working in South Africa, sent a specimen of Euphorbia meloformis, known since 1774 , to the Royal Gardens in Kew near London. At least that’s what he thought. When the plant came into bloom in the succulents greenhouse in 1899, it clearly turned out to be a different species. According to historiography, this unique plant then disappeared from the Kew collection (probably a concealing way of saying that the plant died). Happily, they had made coloured drawings of the flowering plant (fig.1). That gave enough information to Sir Joseph Dalton Hooker, Kew’s resident botanist, to go ahead with a description in 1903.
Fig. 1 The drawing in Curtis’s Botanical Magazine of the first E. obesa in the Western world shortly before it ‘disappeared from the Kew collection’ (read: died).
Euphorbia obesa is a dioecious species, in other words, a plant which develops only female or only male flowers. The drawing clearly shows a female plant, but in a detailed drawing ‘stamens’ are shown (no. 4 on the plate) and in the description it says: “Filaments anantherous, hirsute”, which means that the stamens are bristly hairy (hirsute) and do no bear anther buds (anantherous). It probably concerns rudimentary stamens that do not develop in a female cyathium. Or would Hooker have mistaken the incised slips of the cup-shaped shell (involucrum) for stamens?
E. obesa remained very rare for some time. In 1907, Alwin Berger in his ‘Sukkulente Euphorbien,’ reported that the Kew plant up to then was the only known specimen. And in the part on the Euphorbiaceae in the ‘Flora Capensis’, N.E. Brown in 1915 reported that only female plants were known. In the Netherlands, the species was first admired in 1924 at the first Succulent Plant Exhibition in The Hague. The owner had bought the plant at the World Exhibition held at Wembley in the same year. In 1926 we find the first photo in the Dutch magazine ‘Succulenta’. Because of its rarity, E. obesa was a coveted plant at the time and people were willing to pay large sums of money for it. As a result, the localities were literally looted once they were known. The South African government recognized that there was a threat of extinction and to prevent that, an export ban for live plants was imposed in 1931.
The species name obesa means ‘thick’, ‘fat’, ‘swollen’. Just think of the new health issue obesity, in which the same word is incorporated. In this context, it is worth mentioning that the plant in South Africa is called vetmensie (‘little fat human’).
Another South African name is ‘klipnoors’. Klip means stone or boulder and noors is the name which in the Jansenville area was given to the common yellow-flowered and rather thorny Euphorbias. These reminded the British of the gorse (Ulex europaeus) found in England. This then was probably corrupted by the Dutch to Noors (‘Norwegian’). The region around Jansenville is now called Noorsveld and ‘Noors’ has become a designation for Euphorbias in general. A klipnoors is therefore a Euphorbia that looks like a piece of stone.
Names in English are: Living baseball, Eisenhower’s golf ball, Baseball plant, Sea urchin (zee-egel). This last one obviously as a reference to a sea urchin’s shell. See Fig. 2 and 3 below.
Fig. 2 Euphorbia obesa subsp. obesa, in South Africa called vetmensie (a fatperson).
Fig. 3 The shell of a sea urchin
The species occurs near the town of Kendrew in the Graaff-Reinet district (on the west side of the Eastern Cape). There the plants grow on the top and the southern slopes of low hills, between 300 and 900 meters above sea level and also on the flat parts between the hills. On these flat parts they occur in sandy soil under shrubs, but on the slopes the soil is much more stoney. The colours of the plant body match the environment so well that it is difficult to find one.
In summer, the maximum temperature is on average 26 °C and the minimum temperature 11 °C. In winter it sometimes freezes slightly. The annual rainfall is 200 to 300 mm, spread over two periods. In late spring (October – November) there is some precipitation and at the end of the summer (March – April) larger quantities are registered. Most of the rain falls during thunderstorms. Apparently, the plants are eaten by the numerous baboons and by the cattle of the farmers. The milky juice does not seem to bother these animals.
Fortunately, despite the collecting frenzy of the enthusiasts, there are still some populations left (perhaps overlooked at the time). They are now protected, sometimes even by placing a fence around them.
It is also completely unnecessary nowadays to remove these plants from nature. They are grown in large numbers and the cultivation is certainly not difficult. It is actually incomprehensible that in 1935 it was still thought that the species could hardly be kept alive, as appears from the following fragment from the question and answer section in the October issue of ‘Succulenta’:
“For Euphorbia obesa, the ‘right’ culivation in our country has not yet been discovered. At best, one can keep this most remarkable of all Euphorbias, which only occurs in one part of the Cape Colony (Kendrew in the district of Graaff Reinet northwest of Port Elisabeth), alive for a few years. They slowly wither away in our country: there is usually no question of them getting any bigger.”
For a long time, it was believed that the species indeed only occurs in the immediate vicinity of Kendrew, south of the town of Graaff Reinet, but apparently there are also populations north and northeast of Graaff Reinet.
Sensational was the discovery by Robert Allen Dyer in 1939 of a population of a similar plant on Mr. Stegman’s Kruidfontein farm, 19 miles west of Willowmore, on the road to Rietbron. This is at a distance of more than 100 km southwest of Kendrew and in the intermediate area no plant resembling an E. obesa has ever been found. This plant was described in 1941 by White, Dyer and Sloane as a separate species, E. symmetrica (fig. 4).
Fig. 4 E. obesa subsp. symmetrica.
The main difference with E. obesa, in addition to the geographical separation of course, is that the points from which the inflorescences originate (called flowering eyes), are different in shape. In E. obesa they are round dots from which one flowering stem (peduncle) originates, but in E. symmetrica it is an elongated transverse stripe, and this offers space for several (up to 5) flowering stems next to each other (fig. 5). Incidentally, in E. obesa there may also be several inflorescences together, but in that case, they originate from one branched flowering stem.
Fig. 5 E. obesa subsp. symmetrica differs from ssp. obesa in that the “flowering eyes” are stripe-shaped and can thus accommodate up to 5 flower stalks (peduncles).
In addition, E. symmetrica keeps its spherical shape much longer than E. obesa. In the long run, however, it also becomes taller than wide.
Also, in E. symmetrica the taproot is supposed to develop much more vigorously. Later on it was also mentioned that young seedlings are spherical in E. symmetrica and cylindrical in E. obesa. Gordon Rowley in 1998 found the differences too small to distinguish 2 separate species and he reduced E. symmetrica to a subspecies, E. obesa subsp. symmetrica. The original E. obesa is now called E. obesa subsp. obesa.
In an article by Gerhard Marx in the magazine Cactus & Co, some more differences are mentioned: in subsp. symmetrica the head is more sunken than in subsp. obesa; the colored stripe pattern on the plant body in subsp. symmetrica (especially visible in seedlings) consists of fewer, but wider and more pronounced bands than in subsp. obesa and under identical conditions, especially with regard to the amount of light, the colour of subsp. obesa is more purple blue-grey and of subsp. symmetrica green to yellow-brown. Young seedlings are clearly greener in subsp. symmetrica.
Propagation by seed works very well in E. obesa and the seedlings quickly develop into beautiful little balls. On April 29, 2008, I sowed 25 seeds of subsp. symmetrica (from my own plants) in 5 neat rows of 5 in a pot of 8 x 8 cm (at the top). I always sow at a temperature of 25 °C to 30 °C. Within one week, 13 seelings had already emerged. In fig. 6 we see the pot with seedlings on May 18, so barely 3 weeks after sowing. By then the 22nd seedling had just emerged from the soil.
Fig. 6 E. obesa subsp. symmetrica seedlings 3 weeks after sowing.
In fig. 7 we see the same pot on July 13, almost 11 weeks after sowing. Now there are 23 seedlings on display. A success rate of 92%. If the seedlings are not transplanted, sometimes a seed will germinate in the next year. A late vocation, shall we say.
Fig. 7 E. obesa subsp. symmetrica seedlings 11 weeks after sowing.
As far as the soil is concerned, the plant is not very demanding, but a mineral composition is recommended. Water regularly in the growing period and keep dry in winter. According to the literature, temperatures down to -10 °C. are tolerated when the soil is completely dry.
Subsp. symmetrica is perhaps a little more sensitive to cold. In its habitat it does not get as cold as at the localities of subsp. obesa.
Over the years, a whole series of abnormalities has been reported among the countless cultivated specimens of subsp. obesa. Of course, cristates have been known for a long time. Usually they are grafted but this is not really necessary. One of my seedlings from 1998 changed into the cristate form from the beginning. Fig. 8 shows part of this now 14-year-old plant which still has its own root system.
Fig. 8 A 14-year-old seedling of E. obesa subsp. obesa on its own roots.
E. obesa does not normally form side shoots, but in some specimens, shoots are produced from the base. The designation ‘forma caespitosa‘ was coined for this. Bizarre in appearance is ‘forma prolifera‘, in which a new shoot is created from every growing-point. It is very similar to the witch broom disease that occurs in some cacti.
On to deviations in flowering. Under the not so correct title “Impotence in Euphorbia obesa“, J. Mieras reports in 1978 a male plant whose stamens do not develop and in 1979 a female specimen is reported in which the pistil does not develop normally.
Male plants which become females and vice versa also occur. In addition, in 1983 P.H. den Hartog reported a plant of which all cyathia are bisexual. So, both pistil and stamens in one cyathium. In my own collection I have observed in both subspecies that there are male specimens which regularly produce bisexual cyathia and fruits thereof (fig. 9).
Fig. 9 E. obesa subsp. obesa with some bisexual cyathia.
Furthermore, there is the obesa with deformed stamens mentioned by Riet Maessen in the December issue of Succulenta 2012 (fig. 10).
Fig. 10 E. obesa subsp. obesa with monstrose male inflorescences, photo Riet Maessen.
Then there are the plants that proceed to multiple dichotomous division without this leading to the formation of a crest (cristate). These forms are referred to as ‘forma polytomica‘. Mixed forms of polytomic and cristate growth also occur. Gordon Rowley gives these types of shapes the name “Rocky Mountain”. Unfortunately, the species was not designed to undergo such a drastic change in shape and that means that cracks will occur, resulting in ugly brown spots.
I have a few such forms in my possession, all female ones and there is a lot to experience there. To start with, the pistils are not only 3-fold but also 4-, 5- or 6-fold and after pollination with a normal obesa, this results in fruits with 3 to 6 compartments (fig. 11 and 12).
Fig. 11 A monstrose form (“Rocky Mountain”) of E. obesa subsp. obesa with 3-, 4- and 5-parted fruits.
Fig. 12 Close-up of the fruits in Fig. 11.
On one of those plants, a kind of monstrose way of flowering began to develop 2 years ago in which a still expanding lump of cyathia has arisen (fig. 13).
Fig. 13 Monstrose inflorescence in E. obesa subsp. obesa “Rocky Mountain” .
Fig. 14 On the right a seedling of E. obesa subsp. obesa with 3 seed-leaves.
This is a phenomenon that also occurs quite frequently in cacti. Other seedlings will keep making new ribs so that after 1 year there are already about 13 instead of the usual 8. In the second or third year, such a seedling then proceeds to dichotomous division. In fig. 15 we see such a plant after the division is complete and fig. 16 shows such a plant at a more advanced stage.
Fig. 15 After forming about 20 ribs, this seedling of E. obesa subsp. obesa decided that it was necessary to divide dichotomously.
Fig. 16 A plant of E. obesa subsp. obesa divided dichotomously.
So all this refers to offspring of a monstrose female plant that has been pollinated with pollen from a normal male plant. In any case, the phenomenon seems to have a hereditary component. I wonder how big the percentage of abnormal offspring is if the supplier of the pollen is also a monstrous form.
What remains to be told is the fact that E. obesa hybridises rather easily with other species and that the plants in our collections will not all be true to type. As long ago as 1938, ’Succulenta’ contained a picture of E. obesa x submammillaris. In addition, there is a cultivar on the market under the name of “William Denton” which is claimed to be a hybrid with E. mammillaris. Common are hybrids with E. meloformis. Among other things, these are distinguished from the true obesa by the fact that the flowering stems, just like in E. meloformis, usually are not shed after flowering. Hybrids with E. horrida, E. globosa and E. ferox are also known.
LITERATURE
Berger, A. (1907). Sukkulente Euphorbien. Ulmer Verlag, Stuttgart: 102.
Brown, N.E., Hutchinson, J, Prain, D. (1915). Euphorbiaceae in Thiselton-Dyer, Flora Capensis, William Clowes and Sons, London, Vol. 5, sect. 2, part 2, p. 359.
Hartog, P. H. (1983). De sexuele spelingen van Euphorbia obesa. Succulenta 62 (11): 258.
Hooker, J. (1903). Curtis’ Botanical Magazine 129, tab. 7888.
Houten, van den, J.M. (1926). Merkwaardige Euphorbias, Succulenta 8 (7/8): 100.
Killick, D. J. B. (1978). The flowering plants of Africa.
Laren, van, A, J. (1932). Vetplanten, Verkade’s fabrieken N.V., Zaandam.
Marx, G. (2012). The South African spherical spurges, Cactus & Co 16 (2): 70 – 81.
Mieras, J. (1978). Impotentie bij Euphorbia obesa, Succulenta 57 (7): 150.
Mieras, J. (1979). Nogmaals Euphorbia obesa, Succulenta 58 (8): 203.
Rowley, G. (2006). Teratopia, Cactus & Co.
Sluys, van der, C. (1938). Euphorbia obesa x Euphorbia submammillaris, Succulenta 20 (1): 11.
Thoorn, J.J.E. van de, (1935). Vragenrubriek, Succulenta 17 (10): 159.
White, A., Dyer, R., Sloane, B. (1941). The Succulent Euphorbieae 2: 559, Appendix A: 964.
First published in Succulenta 92 (6), December 2013. Translation from the Dutch by F.N.
C. columnaris was described as early as 1778 by the Swedish physician and botanist Carl Peter Thunberg. He has been called ‘the father of South African botany’ and ‘the Japanese Linnaeus’. From 1772 to March 1775 he was in South Africa and it is certain that in September 1772 he collected plants in the Little Karoo, one of the areas where this species occurs. From 1775 to 1778 he was a medical doctor in Deshima, the Dutch settlement in Japan. On his way to Amsterdam, he spent another 2 weeks in South Africa in 1778. It is plausible that Thunberg collected the plant himself or at least saw it in its natural habitat and therefore it surprises me that he gave it the name columnaris (= columnar).
In nature, C. columnaris is almost spherical, flat rather than elongated. It has been noted several times in literature that the species better could have been named C. globosa (= spherical). Perhaps Thunberg continued to cultivate the plants that he took with him for a while, resulting in them losing their natural appearance. This is a problem that these types of plants tend to suffer from in cultivation.
English-speaking people often use the name ‘khaki button’ which, in addition to the shape that the plant assumes in the dry season, also refers to the distinctive color that the plants adopt (see fig. 1).
The Afrikaans name ‘koesnaatjie’ for this plant also refers to the flattened convex appearance. A naatjie, also called naartjie, nartjie or nartje is a small citrus fruit that is halfway between a mandarin and a tangerine. Koes means in hiding, withdrawn. A mandarin in hiding more or less, probably referring to the way in which the plant partially withdraws into the ground in the dry season (I assume that the plant does not taste like a mandarin, even if it is edible). On the internet I found the following description of the koesnaatjie: “A koesnaatjie is a succulent plant found in the dry western and southwestern regions of the Cape. The plant has thick, fleshy, oval-shaped leaves which are fused at the base and fold tightly over each other like roof-tiles, so that the plant has a compact appearance, and it is edible. Only once does it develop an inflorescence with densely congregated cream to yellow flowers after which the plant sets seed, withers and dies”.
In the 20’s and 30’s of last century, the Dutch monthly magazine ‘Succulenta’ regularly wrote extensively about this plant and the differences between nature and cultivation were always pointed out in detail. For example, in the November 1935 issue I read:
Miss W. v. A. in B. writes: “Can I take cuttings of an etiolated Crassula columnaris? They are so ugly with that brown piece of stem. What causes these plants to grow so spindly here?”
And the editors’ answer:
“The proper cultivation of highly succulent Crassula species in our climate is not easy. Many of these plants originate from the Karroo highlands. The average rainfall there is 4.47 inch ( ± 114 mm). In our country, an average of 700-800 mm rainwater falls per year. The rain on the Karroo plain falls mainly in the four winter months, sometimes also in November and December. These two months of summer rain have little influence on growth, but are important for replenishing the water reservoirs of the succulents.
Few deserts have a more desolate appearance in the dry period than the Karroo. There are no green plants to be found, the gullies are dry and the isolated “Koppies” (small mountain peaks) which are scattered here and there in the Central Karroo, seem to reflect the heat like a mirror. During the night a cool S.E. breeze prevails. The imaginary line which divides the districts into regions with summer or winter rains, runs through the Central Karroo near Beaufort West. In some winter nights the temperature drops below freezing point, and the mountain peaks are sometimes covered with snow, which, however, rarely falls on the plateau itself. The average daily temperature difference is large, i.e. 30° F (= 17 °C). Due to the intensely dry air, the summer heat is not very oppressive. In the Northern Karroo the average rainfall per year is even less, in the Western part 2 inches, so 51 mm per year. The average summer temperature in the shade is 110° F (= 43 °C). This gives us an idea of the conditions under which the highly succulent Crassulas grow in their homeland and makes us understand that it is therefore not easy for these plants to maintain their beautiful, stocky appearance here. Crassula columnaris occurs near Whitehill, the first train stop past Matjesfontein. Jacobsen mentions Namaqualand as habitat. The annual rainfall there is about 6 inches = ± 150 mm. Crassula columnaris allows you to take cuttings (the best time for this is August because the plant starts to grow in this month. After rooting, be very, very economical with water. The entire structure of the plant indicates moisture absorption from the air. In summer provide as much sun and fresh air as possible; the plants may turn reddish brown.”
It is remarkable that between 1923 and 1942, 5 times was written extensively about C columnaris in ‘Succulenta’ and after that, which is now over 70 years, nothing at all. The name is only mentioned in passing once in a while.
As can be seen from the above, C. columnaris belongs to the group of winter growers, which have their resting period in summer and should receive minimal water then. In many books a winter temperature of at least 15 °C is recommended, but in my experience that is not good at all. Such a high temperature combined with the application of water promotes growth and with the short days and the low light intensity in winter, this means an elongated, unnatural shape. I have never noticed that the temperature in my greenhouse – which I try to keep at least 8 °C in the winter (with varying success) – would harm the well-being or flowering of such plants.
Other members of the group of winter growers are C. alstonii, barbata, deceptor and pyramidalis. They are best grown in sandy soil with a small proportion of humus.
C. columnaris is monocarp, which means that the plant dies after flowering. This prompted Gordon Rowley in his book ‘Crassula’ to describe cultivating C. columnaris as “a passing pleasure”. Beautiful plants of the species are rare in our collections. Seedlings become elongated and imported plants immediately bloom and then die. To keep the plants somewhat presentable, Rowley recommends a starvation diet with plenty of sun and fresh air.
Although the first signs of flowering also announce the impending death of the plant, it is still fascinating to see how the inflorescence develops. It starts with the closed rosette bursting open at the top, as it were, to provide room for a compact arrangement of flower buds. See Fig. 2.
Fig. 2 The flower buds burst out of the plant, as it were
This is a plant I bought as a young seedling with the name “C. columnaris Lemoenshoek” at the Special Plant Market in Nijmegen. Lemoenshoek is a farm about 40 km east of the village of Barrydale in the Little Karoo. The photo was taken on December 3, 2012. The diameter of the plant in the photo is 4 cm and its height about 3 cm. In those 3 cm, 7 pairs of leaves are pressed together. Once in full bloom, the plant body is completely hidden from view by the numerous white flowers. See the photo of Fig. 3, which was taken on January 19, 2013. The name ‘shaving brush crassula’ used in English-speaking countries needs no further explanation.
Currently, 2 subspecies of C. columnaris are distinguished: in addition to C. columnaris ssp. columnaris (the standard form that this article is mainly about) there is also C. columnaris ssp. prolifera. The description by H.C. Friedrich dates from 1974. This subspecies branches off at the base and the inflorescence is not right above the rosette as with the other subspecies. When ssp. prolifera is going to bloom, the central axis lengthens so that the plant approximately doubles in height. At the top of the central axis, which is loosely covered with small leaves, the inflorescence develops, which is also less compact than in the other subspecies (see Fig. 4 below, photo taken on the Knersvlakte in August 2004 by Sofia Etzold), with thanks to ‘Photo Guide to Plants of Southern Africa).
In Afrikaans, this variety is called ‘sentkannetjie’. ‘Sent’ in this expression is derived from the English ‘scent’ and kannetjie stands for bottle. This does justice to both the shape of the plant and the sweet smell of the flowers.
Hybrids of C. columnaris are uncommon, but H. R. Toelken in the Flora of Southern Africa mentions a putative natural hybrid C. alpestris ssp. massonii x C. columnaris ssp. columnaris and gives the following information:
“The plants superficially resemble C. columnaris subsp. columnaris with its clasping leaves producing an erect column c. 15 mm in diameter. However, the plants are longer than broad and have terminal and axillary inflorescences and the leaves are bluntly acute as in C. alpestris ssp. massonii. The putative parents have been recorded from the area”.
Finally: C. columnaris should not be confused with C. columella, which has many similarities with C. columnaris in both name and appearance. This plant forms thin, branching, square columns of up to 1.5 cm thick and 5 cm high (columella means ‘small column’), consisting of tightly packed (tile-like) leaves. See below.
Fig. 5 Crassula columella also flowers terminally but does not die afterwards. Next to the faded head (right), one or two new shoots arise.
It also flowers terminally, with small, inconspicuously colored flowers on a stalk. Unlike C. columnaris, the stem does not die after flowering but forms 1 or more side shoots directly next to the flower stem, so that a low bushy plant is developed. For us, plant fanciers, this has the great advantage that the plant not only becomes more beautiful after flowering, but also provides a lot of easily rooting cuttings. This species is also hardly prone to the etiolation that C. columnaris so easily suffers from.
Literature:
Laren, van, A,J. (1931). Succulents, Verkade’s factories N.V., Zaandam.
Rowley, C. (2003): Crassula, Cactus & Co.
Swüste, F (1935). Vragenrubriek, Succulenta 17 (11) p. 175.
Thunberg C.P. (1778). Nova acta physico-medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum.
Photo Guide to Plants of Southern Africa, www.southernafricanplants.net
First published in Succulenta 92:5, October 2013. Translated from the Dutch by FN.
For more habitat pictures and other info follow this link: Crassula columnaris (part 1 of 3)
Guest column by Theo Heijnsdijk (Translation and habitat pictures by FN)
History In 1812 Adrian Haworth in his “Synopsis plantarum succulentarum” described a new genus in the family Euphorbiaceae: Dactylanthes, translated by himself as Finger-flower. He distinguished four species in the genus: D. patula, D. anacantha, D. tuberculata and D. hamata, characterized by the finger-like lobes of the inflorescences.
In 1823 he added a fifth species: D. globosa. In the description, he reported that he had seen the species in bloom in Kew in October of that year.
Fig. 1
In 1826, Curtis’s Botanical Magazine published a coloured plate (plate 2624, see figure 1) with a detailed description by John Sims. This time under the name Euphorbia globosa, which is now generally recognized. The genus Dactylanthes has thus merged into the genus Euphorbia. The species D. anacantha and patula are seen as synonyms of Euphorbia tridentata by current botanists. For the other three, the specific name has remained the same.
Habitat This distinctive plant seems to have been widely distributed in the Eastern Cape area of South Africa*, but nowadays the species is much rarer. It occurs in the area between Port Elizabeth and Uitenhage where it grows on low stony hills up to 20 km from the coast.
On the Red List of South African plants, the species has the status ‘endangered’. According to that report, it is known from less than 5 localities. The plant is listed in Appendix 2 of CITES.
* In the famous work “The succulent Euphorbieae” (1941), by White, Dyer and Sloane, we find (among many other things) the following observation on this topic: “E. globosa is abundant in the karoid scrub between Uitenhage and Port Elizabeth, sometimes forming uniform mats of the dwarf branch-tips in such masses that it is difficult for the passer-by not to tread on them.”
Description
It forms compact clumps up to 8 cm high and to 50 cm in diameter, consisting of globular** stem segments that seem to be connected in a rather random way.
** Hence the name globosa (= spherical).
The young shoots bear small scaly leaves which later disappear. The oldest segments are grey to almost white.
African names for the species are eierpol, knopmelkbol and langbeentjie.
A mature segment develops a thin stem that bears the inflorescence, which is supported by 3 bracts. When the inflorescence or fruit has dropped, the flowering stem often divides again into 1, 2, or 3 new stems which like the older ones may become about 8 cm long.
Fig. 2
Fig. 2 shows the scar of the dropped inflorescence, and it is also visible how 2 new stems develop alongside. At the bottom, the third one just starts. A trichotomous division I would say. In this way, a cascade of stems is created. The dried flower stalks remain on the plant. White, Dyer and Sloane have the following comment on the species:“Much has been witten about E. globosa since 1826, yet its “singularly whimsical growing habit” remains as fresh a source of surprise to-day as ever. In its native surroundings the globose branches form thick mats, closely pressed to the ground. These branches are gradually drawn down almost perpendicularly underground, new branches replacing them at the surface, and these take their turn for a while at putting forth the characteristic cyathia and cymes, on peduncles which are variably very short or almost absurdly long. Under cultivation the branches are rarely drawn underground, but they pile up in a fantastic medley, each branch differing somewhat from the others in shape, globose, clavate or cylindric, so that a large potted plant looks not unlike a bowl of quaint, irregular marbles or tangled strings of misshapen large beads. Occasionally the longer peduncles produce little new branches up aloft, instead of the expected inflorescence, until of their own weight these adventitious newcomers drop over to the ground, send out rootlets and begin life on their own accord.”
Inflorescence
Here I would first like to discuss the special inflorescence of Euphorbias. What we see as a flower is in fact a whole bouquet. Each stamen is a separate male flower stripped of all non-essential parts. Only the stamen itself, ending in 2 anthers, is therefore left. In the same way, the pistil is in itself a separate female flower. Pistil and stamens grow from a kind of cup, called involucrum. On the outside of the involucrum there are usually 4 or 5 nectar glands. All those basic flowers, together with what is around them, form what an ordinary plant enthusiast sees as ‘the flower’. But in botany, this combination of flowers is called a cyathium.
Fig. 3
In figure 3 we see a young stage of the very special inflorescence of E. globosa. In the middle you can see the pistil with the 3 stigma lobes and around it the anthers in an immature stage. The diameter of this inflorescence at that time was 10 mm. If you look at that picture you will immediately understand why Haworth chose the name ‘Dactylanthes’ (finger-flower) for this type of plant.
The ‘fingers’ are outgrowths of the nectar glands. The nectar is secreted by the wart-like elevations. In E. globosa we usually find 3 or 4, sometimes 5 ‘fingers’ per nectar gland.
Fig. 4
In figure 4 we see an inflorescence at a much later stage. The pistil has reached its final size and the stigma with its 3 lobes sticks out between 2 fingers. The swollen ovary is clearly visible. At the bottom left of the cup we see 2 immature anthers and at the top right a stamen with 2 ripe, pollen producing anthers. If you look closely, you can see about halfway up this stamen, at the level of the cup edge, a kind of ring around the stamen. That is the transition from flower stalk to stamen. When the stamen (= flower) has finished flowering, it drops off its flower stalk. What remains is a kind of hollow tube. In the cup we also see a number of flower stalks from which the finished stamen has already broken off. On the 3 fingers of the honey gland at the bottom right, we see a few of those broken stamens.
A peculiar thing in the photo is that at the upper nectar gland there were first 3 fingers of which the middle one has divided again halfway.
It is reported that at the ends of the flowering stems sometimes not cyathia, but small spherical sections develop. If the stems bend due to the weight of the segments and touch the soil, they will quickly take root.
In cultivation I bought my oldest E. globosa around 1990 at a cactus market. It was probably already a slightly older plant, consisting of some larger gray-white balls, and I wondered if it had been imported. This plant has now grown into a collection of perhaps 70 stems, covered with a tangle of dried flowering branches which makes it impossible to make a nice photo of it. This plant is self-fertile. Usually, after flowering, the fruits with 3 fertile seeds -characteristic for Euphorbias- appear .
Fig. 5, a seedling of a few years old
In figure 6, we see seedlings of 5 months old in a pot.
In figure 7, photographed when the same seedlings were 4 months old, it can be seen that one of the plants in the same pot is already preparing to flower.
Euphorbia globosa requires a sunny place with not too much water. If not, the spheres lengthen, and the plant will look like the closely related E. ornithopus.(The same group also includes E. tridentata, E. polycephala, E. wilmaniae and E. planiceps).
Minimum temperature in winter 8 ºC, although in literature it is reported that the plant can withstand frost.
Finally In a series of articles about the plants grown in the botanical garden in Geneva, Alphonse de Candolle wrote an extensive article about E. globosa in 1836. There was a beautiful folding plate in which all parts of the plant were depicted in detail. See below. Unfortunately, the scan is slightly distorted on the left-hand side.
Literature Buddensiek, V. (1998). Sukkulente Euphorbien; 25, 101.
Candolle, A. P. de. (1836). Septième Notice sur les Plantes rares cultivées dans le Jardin de Genève, Memoires de la Société de physique et d’histoire naturelle de Genève 7 (2): 288 – 293, fig. 5.
Haworth, A. H. (1812). Synopsis plantarum succulentarum: 132.
Haworth, A. H. (1823). Plantae rarae Succulentae; a Description of some rare Succulent Plants, The Philosophical Magazine and Journal 62: 382.
Sims, J. (1926). Euphorbia globosa, Curtis’s Botanical Magazine 53, plaat 2624.
White, A; Dyer, R.A. and Sloane, B.L. (1941). The succulent Euphorbieae (Southern Africa).
“The most magnificent and beautiful of all the Crassulaceae is the scarlet Rochea coccinea. Emerging from crevices or growing freely on the flat rocks, the large perennials accommodate their root system in a layer of soil only a few centimeters thick, which sits so loosely on the rock that it can be lifted off together with the plant. The fact that despite its fleshy leaves it is dependent on the moisture coming from the summer clouds is best demonstrated by the cultivation experiment in Cape Town: it only thrives there, even moved to similar locations, if it is watered at least once in a while in summer.”
This is a quote from the book ‘Das Kapland’ by Rudolf Marloth, published in 1908.
According to literature, C. coccinea grows at altitudes over 800 meters in the Southwest Cape. As distribution area, the Cape Peninsula (especially Table Mountain near Cape Town) and the area between Paarl and Bredasdorp (135 km to the southeast) are mentioned.
In reality the area is considerably larger, as evidenced by the photo taken by Bertus Spee in mid-November 2008 in Citrusdal, 125 km north of Paarl. This was at the beginning of the flowering period. In nature it is part of the so-called fynbos vegetation (Mediterranean-like vegetation).
Fig. 1 C. coccinea and an Adromischus in 2008 at the beginning of the flowering period (mid-November) in Citrusdal (Western Cape). Photo Bertus Spee.
The stems become about 40 cm high and they branch from the base. In older plants, the stems become bare and brown from below, with only the tips bearing leaves. Flowering occurs in summer (in Europe in winter). Due to the long flower tube of about 4 cm, only butterflies can access the nectar. Back in 1925 Marloth noted that the butterfly Meneris (now Aeropetes) tulbaghia is the main pollinator. This butterfly is called “the table mountain beauty” or “the mountain pride butterfly” in South Africa and is specialized in red flowers. It is rumoured that you will be chased by the butterfly if you wear a red cap. It is probably the only pollinator for at least 15 plant species in fynbos. Among those 15 are 3 species of the genus Cyrtanthus, which is also known to succulent lovers, and 7 of Gladiolus.
History Like Kumara (Aloe) plicatilis discussed in an earlier post, Crassula coccinea belongs to the succulents that were in cultivation in the Netherlands as early as the late seventeenth century. It is first mentioned in a book from 1706 by Caspar Commelin about rare plants in the Hortus Medicus, the predecessor of the current Hortus Botanicus in Amsterdam. The ‘name’ that Commelin gave was ‘Cotyledon Africana frutescens flore umbellato Coccineo’ which means something like: bushy growing African Cotyledon with scarlet flowers in a flat-topped inflorescence. The accompanying black-and-white drawing shows a plant with flower buds about to open.
Fig. 2 Image of Crassula coccinea in Casper Commelin’s 1706 book.
In the Moninckx atlas (see also the earlier post about Kumara plicatilis) there is also a beautiful colour image. According to the University of Amsterdam, this was made somewhere between 1686 and 1706 by Maria Moninckx. Although the images clearly differ from each other, they both seem to show the same specimen.
Fig . 3 Watercolour of Crassula coccinea by Maria Moninckx (made between 1686 and 1706).
The image in the book ‘Phytanthoza iconographia’ by Johann Wilhelm Weinman from 1737 is also beautiful. Here C. coccinea is depicted together with C. perfoliata and C. tetragona. The funny thing is that a mistake was made with the Latin specific name. It says “fructescens” (bearing fruit) instead of “frutescens” (shrub-forming). Not just a clerical error, because in the German text the plant is called “Fruchtbringend”.
Fig. 4 Image of Crassula coccinea (a), C. perfoliata (b) and C. tetragona (c), in Phytanthoza iconographia from 1739.
Nomenclature
As far as I can tell, the name Crassula first appears in 1732. The German botanist Johann Jakob Dillenius used it for eight succulents in his work ‘Hortus Elthamensis’. The name means something like ‘fat one’. The common name is ‘thick leaf’. Linnaeus adopted the genus name in his ‘Genera Plantarum’ in 1735 and in 1753 he already described 10 species of Crassula in his ‘Species Plantarum’ (first edition). It’s curious that he mentions Ethiopia as the place of origin for Crassula coccinea, as well as for 7 other Crassulas. Of the 2 remaining ones, C. alternifolia is only given a question mark and in the case of C. scabra, he mentions “Habitat in Africa?”.
Later on, C. coccinea was placed in the genus Rochea. A.P. de Candolle created this genus in 1802 to accommodate the Crassulas with large tubular flowers. The genus name refers to the Swiss botanist Daniel de la Roche. In 1805 the South African mycologist Christiaan Hendrik Persoon changed the genus name to Larochea, after which the Austrian Leopold Trattinnick (botanist and also mycologist) changed it into Dietrichia in 1812. Adrian Haworth in his turn created the genus Kalosanthes in 1821 to accommodate 8 Crassulas including C. coccinea.
He also reported a white-flowered variety (“floribus albis”) based on a book with drawings and descriptions of succulent plants by Richard Bradley from 1716. Unfortunately, a number of drawings in that book were incorrectly numbered and that resulted in, among other things, C. coccinea containing the text of C. tetragona. And there was talk of “small white flowers”. One wonders how well Haworth looked at his sources, as C. coccinea has the largest flowers of the entire Crassula genus.
All the above genus names have been declared invalid and it is again just Crassula. Kalosanthes has remained as the name for one of the 20 sections within the genus Crassula. This section contains only 3 of the over 200 Crassula species. Apart from C. coccinea they are C. fascicularis (= C. odoratissima) and C. obtusa (= C. jasminea). See also fig. 5, where the species of the section are depicted.
Fig. 5 In addition to Tylecodon reticulatus, the 3 Crassulas from the current Kalosanthes section are also pictured. Way back in 1912 Marloth indicated that C. coccinea for its pollination depends on the butterfly Menera (Aeropetes) tulbaghia.
Characteristic of the plants in this section are the long tubular flowers which flare out saucer-shaped at the end. This shape is caused by the fact that the 5 petals are fused over at least half their length and have a widened end at right angles.
In South Africa, C. coccinea is called ‘klipblom’ (stone flower). Also common is the name “Red Crassula” and in older literature one may come across the name ’emperor’s crown’.
Cultivation C. coccinea was once a widely cultivated indoor plant. In Victorian times, the plant was loved in England for its large bright red flowers and it is one of the few succulents that has been grown commercially on a large scale . The other 2 crassulas from the Kalosanthes section were also widely grown in England at that time. C. obtusa under the name Rochea jasminea -referring to the jasmine-like white flowers and C. fascicularis under the name Rochea odoratissima because of the strong scent.
In Germany too people were involved in growing and breeding this species. Gordon Rowley in his book ‘Crassula, A Grower’s Guide’ mentions that the famous Haage firm in Erfurt offered three cultivars in their 1857 catalogue. In 1868 there were already ten. They have all disappeared from the collections again.
Among the succulent plant lovers we don’t see C. coccinea very much anymore. Young plants are still fresh and compact, but soon they become too tall and untidy. Nevertheless, the species is still grown professionally.
It grows easily and quickly in well-drained soil. Often a mixture of two parts of leaf mould, one part clay and one part sand is recommended. In full sun, a lot of water and fertilizer is required. The plants can be grown from seed or cuttings. Seedlings can already bloom in the first year. The plants are susceptible to the fungal disease ‘rust’ but can’t stand fungicides (pesticides against fungi). After a few years, the plants become ugly with dying leaves at the bottom and deflecting shoots. That is the time to take cuttings. In a mixture of peat and sand they will quickly root.
Cuttings can also just be put in a jar of water. Once they are growing, heading them several times is recommended. This creates compact bushy plants. Supplying a larger pot may also be needed regularly. In summer they can stay outside without problems. In winter keep them cool but do not let them dry out.
Fig. 6 Crassula coccinea flowering in cultivation. The species was massively grown in the 18th century for its large, bright red flowers.
Hybrids In comparison to some other Crassulas, C. coccinea has not often been used for hybridisation. Probably the oldest hybrid is C. coccinea x falcata. This is midway in appearance between the two species and was named Kalorochea langleyensis. In1898, a specimen was exhibited at the Royal Horticultural Society’s show in Chelsea. This hybrid, which according to the current rules should be called Crassula x langleyensis does not seem to be cultivated anymore.
A hybrid with C. undulata was also once produced, but has suffered the same fate and the same applies to the cross with the white-flowering C. obtusa (Rochea jasminea).
Literature Arianoutsou-Faraggitaki, M & Groves, R, H. (2012). Plant-animal interactions in Mediterranean-type ecosystems: 137 – 147.
Candolle, A.P. de & Redouté, P.J. (1799-1837): Plantarum Historia Succulentarum, 1: 1.
Commelin, C. (1706). Horti medici amstelædamensis plantæ rariores et exoticæ ad vivum æri incisæ: 24.
Dillenius, J. (1732): Hortus Elthamensis 1: 114 – 121.
Knippels, P.J.M. (1992). The household consumption and turnover of succulents at the flower auctions, Succulenta 71 (1): 33 -38.
Linnaeus, C. (1737). Genera Plantarum: 89.
Linnaeus, C. (1753). Species Plantarum 1: 282 -283.
Marloth, R. (1908). Das Kapland, Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer “VaIdivia” 1898-1899, bd.2, T3:142, 339.
Marloth, R. (1925). The Flora of South-Africa 2: 17 – 24.
Kalanchoe delagoensis is sometimes regarded in quite different ways. Those who Google Kalanchoe tubiflora (the name under which the plant is better known), will find both “How to grow Kalanchoe tubiflora” and “How to kill Kalanchoe tubiflora”. The latter is not intended for the many plant lovers who consider this plant to be weeds in their greenhouses, but for use in the many areas of the world where the plant has started to grow and poses a threat to livestock because of its toxicity. More on this later in this article.
There are also differences of opinion about what is the correct name for this plant.
For starters, Bryophyllum is very often chosen as the genus name. This name aptly reflects the property of the plants in question to produce miniature plants on the margin of the leaves that detach easily and then quickly develop into powerfully growing plants. Others choose to consider Bryophyllum as a section in the genus Kalanchoe.
The origin of the name of the genus Kalanchoe (described by the French botanist Michel Adanson in 1783) is not at all clear. Maybe it’s a phonetic transcription of the Chinese jia lan cai shu (I also came across kalan chau huy) and this would have been corrupted to Kalanchoe. Jia lan cai shu is said to be the name of a species occurring in China, possibly K. ceratophylla or K. spathulata. Searching the internet, I found out that nowadays the Chinese name for Kalanchoe is also Jia lan cai shu(see Flora of China 8: 204–205. 2001), so I wonder if it is not the other way around, to wit that the Chinese name is a corruption of the botanical name.
Other sources refer to the names from Sanskrit kalańka-, “stain” or ” rust”, and chāyā, “clear” which would refer to the leaves of the Indian species K. laciniata.
I also read somewhere that kalan chau huy would mean “that which falls and grows” which would be a perfect description of the plants of the Bryophyllum section. But this seems like “wishful thinking.” All species of the Bryophyllum section occur naturally exclusively in Madagascar which is very far away from China.
With regard to the species name, it should first be noted that “delagoensis” refers to Delagoa Bay (now Maputo Bay) near the capital Maputo of Mozambique (until 1975, when Mozambique became independent from Portugal, called Lourenço Marques). A surprising name for a plant that originates from Madagascar which is located some 1200 km from Delagoa. In order to understand the discussion properly, it is necessary to look into history.
It started in February 1822, when the British royal ship, the “Leven” (equipped with 28 cannons) under the command of Captain William Fitzwilliam Owen, together with the accompanying ship the Barracouta (10 cannons), embarked on a 5-year expedition to map the entire east coast of Africa as well as Madagascar and other islands in the area. The entire expedition is described in detail in a report published in 1833 (2 books, together almost 850 pages) by Captain Owen. Two years later, a similar but even more extensive report, by Second Lieutenant Thomas Boteler (2 books, 900 pages) was published. Onboard the Leven was the young botanist John Forbes, dispatched by the British Horticultural Society. Via Lisbon, Madeira, Tenerife, the Cape Verde Islands, Trinidad, Rio de Janeiro, and the Cape of Good Hope, the ship reached Delagoa Bay for the first time at the end of September of the same year. In Rio de Janeiro alone, the tireless Forbes had already collected more than 300 plants for his herbarium, in addition to numerous seeds and rhizomes.
In later years Delagoa became a kind of base from which all kinds of expeditions were undertaken. It was regularly reported that the many mosquitoes kept the people who were ashore awake. More and more people also became ill. There was no knowledge of malaria and the only treatment known against the mysterious fever was bloodletting.
When no less than 19 of the 60 crew members had died at the beginning of December 1822, Captain Owen decided to head for Madagascar. On December 5, the ship departed and on December 21 they arrived at St. Mary’s Island (Île Sainte-Marie) on the eastern side of Madagascar. On January 8, the Leven left for the much more northerly Comoros, and sailing along the coast of Mozambique they were back in Delagoa Bay at the beginning of March 1823. During a stay in Cape Town in May 1823, Lieutenant Browne, botanist Forbes, and assistant physician Kilpatrick volunteered for an expedition upstream from the mouth of the Zambezi River. The intention was to go upstream for some 600 miles and then try to cross to the Cape of Good Hope. With a large canoe and 2 ‘black’ helpers, they entered the estuary on July 23. Forbes fell ill on August 3. Several bloodlettings were to no avail and on August 16 he died, aged 25. Soon after, both of his comrades fell ill too. Browne died on September 5 and Kilpatrick on October 28. The two helpers were also sick but survived.
Captain Owen, meanwhile, was already at Delagoa, where he raised the British flag on the south coast in September. At the bay 4 rivers flow into the sea and he realized the importance of this large natural harbour. Until 1875, this was discussed with the Portuguese who finally got the better end of the stick.
We now jump to the years 1835 – 1837. Then the botanist duo Ecklon & Zeyher (the Dane Christian Friedrich Ecklon and the German Karl Ludwig Philipp Zeyher) published a three-volume book on the vegetable world of southern Africa entitled ‘Enumeratio Plantarum Africae Australis Extratropicae’. In part 3 (1837) we find the following ‘description’:
“KALANCHOE delagoensis. — Exemplum unicum et mutilum Cel. Commodore Owen ad „Delagoabay” legit et nobiscum communicavit. Flor. Jun. — Flores saturate rosei JUN. 1836”.
Mind you, this is the full text. The actual description only consists of the 3 words: Flores saturate rosei. In any case, it can be seen from the text that there was only one mutilated (damaged) specimen and that it ended up at Ecklon &Zeyher via Captain Owen from Delagoabay. I then assume that this happened after Forbes died in August 1823.
For me, the question now is when and where botanist Forbes found the plant. Like all Kalanchoes, K. delagoensis is a short-day plant and it blooms in winter. In the southern hemisphere that’s the case between May and October. Forbes was first in Delagoa from early September to December 5. If the plant had been in bloom at the time, why would botanist Forbes have kept that cutting on board for more than six months? He had a habit of sending all his planting material to England. After the first stay, Forbes was not back in Delagoa until early March 1823. Then the plant is probably not yet in bloom. So how could he have found a flowering plant in Delagoa? Maybe he didn’t find it until July when he was preparing for the expedition upstream along the Zambezi River?
Forbes’ notes may provide more clarification about the find, but they are lodged with the Horticultural Society and are not available on the internet.
In 1862, 25 years after the description of Ecklon & Zeyher, the Irish botanist William Henry Harvey (1811-1866) in the Flora Capensis published the same plant under the name Bryophyllum tubiflorum. Harvey knew the earlier description of Ecklon & Zeyher (he also worked with Ecklon), but according to him this description was invalid and Kalanchoe delagoensis was therefore a so-called nomen nudum, a name without a valid description. Harvey not only chose to place the plant in the genus Bryophyllum because the flowers have only 4 petals, but he also changed the species name delagoensis to tubiflorum. Thinking that the plant should have a different genus name may be understandable, but changing the species name without a proper reason is, in my opinion, very disrespectful to your predecessors. The following is his description: B. tubiflorum (Harv.) ; leaves unknown; corolla thrice or four times as long as the sharply 4-cleft calyx, its segments broadly oblong, very blunt or truncate ; stamens as long as the tube of the corolla. Kalanchoe Delagoensis, E. $ Z.! 1955. HAB. Delagoa Bay, Forbes! (Herb. Sond.) Of this very remarkable plant a portion of a denuded branch, and a part of a dense, probably thyrsoid, inflorescence exist in Herb. Sonder. The internodes are scarcely an inch long, and there are 4 cicatrices, indicating whorled leaves, at each node. calyx 3 lines long. corolla uncial, bright red, its lobes almost square, 2½ lines long.
This description shows that Harvey described exactly the same damaged specimen as Ecklon &Zeyher. We are also briefed on the nature of the damage: the material consists only of a defoliated stem (“leaves unknown”) and part of the inflorescence. If Harvey had seen the tubular leaves, he might have called the plant B. tubifolia instead of B. tubiflora. Tubular flowers are not uncommon in kalanchoe-like plants, but tubular leaves may be unique. For the truly curious among the readers: “Herb. Sond”. in the description above refers to the herbarium of the German botanist Otto Wilhelm Sonder, co-author of the Flora Capensis.
More than a century later, in 1985, Toelken argued that the 3 words of Ecklon & Zeyher were enough to distinguish the plant from its closest relatives so that, according to him, because the name K. delagoensis was the first published one, it should be considered the valid one, rendering the name B. tubiflorum of Harvey invalid. It’s called a nomen invalidum.
There is also Kalanchoe verticillata, described by George Francis Scott-Elliot in 1891 (later renamed Bryophyllum verticillatum by Alwin Berger). According to current views, this is synonymous with Kalanchoe delagoensis.
The stems of K. delagoensis become up to 1.2 m tall and the cylindrical leaves are 1.5 to 12 cm long and 2-6 mm in diameter. The number of teeth at the tip of the leaves is 2 to 9. The terminal inflorescence reaches a height of 30 cm. The bell-shaped flowers are brick-red to reddish violet-gray and 2 to 3 cm long.
Kalanchoes are short-day plants, and the flower-stalk begins to develop around mid-November in my greenhouse.
The plant puts all it has into this flowering and deteriorates dramatically during it. From the top of the plant down, the leaves are ‘drained’ and they start drooping more and more. By the time the plant has finished flowering -this can take some five weeks- there is only little left of it. But, in that pathetic little pile there are still many adventitious plantlets which under favorable conditions will quickly develop into new plants.
With this plant, it seems completely unnecessary to discuss its growing conditions. Nevertheless, I would like to point out that you get the most beautifully developed plants by giving it a lot of light (but not sun all day). It is also not wise to let the plant dry out. It will survive, but it’s definitely not going to get any prettier. It seems that the plant can tolerate a bit of frost. I never tried it.
As mentioned above, the Bryophyllums come from Madagascar and in particular the southwest. F. Vandenbroeck describes in Succulenta that the plant grows noticeably more poorly and stockier in Madagascar’s natural biotopes than in our collections. There were hardly any adventitious plantlets to be seen and the plants did not grow rank. Moreover, the flower colour was much brighter than with us.
Thanks to its phenomenal power of reproduction, K delagoensis has spread all over the world. For example, in the Canary Islands, where I have often found the plant escaped from nearby gardens and from the craving of the garden owners to regulate, much like Crassula multicava discussed earlier in this series.
In many countries, the plant is on the list of weeds. The plant became established in the Australian states of New South Wales and Queensland around 1940. As a short-day plant, K. delagoensis also blooms there in winter (from May to October) which is the dry season and therefore also a time of food shortages. As a result, the cattle often feast upon this very plant, which can result in deadly poisoning. (For the specialists: the poison belongs to the bufadienolide cardiac glycosides). The animals die within 1 to 5 days. In New South Wales, for example, 125 animals died after eating this Kalanchoe in 1997. Rapid intervention by a veterinarian can save the cattle, but that is a pricey issue. Medication for 1 cow costs $70 and this does not include the fee for the vet. That is why no mean measures have been taken.
The legal requirement is: “The plant must be fully and continuously suppressed and destroyed and the plant must not be sold, propagated or knowingly distributed”.
Owning or selling the plant is an offense unless you have a permit. The same measures apply to the hybrid K. daigremontiana x delagoensis (Kalanchoe x houghtonii). This hybrid is very similar to the mother plant K. daigremontiana, but the leaves are much narrower, about 1 cm rather than 4 cm. A characteristic of this hybrid is the sharp incision in the middle of the leaf.
Pest control is done with herbicides and also organically with the South African citrusthrips. A weevil, Alcidodes sedi, is also sometimes used for biological control.
For a plant that is spread worldwide, of course, many names are in circulation. In English e.g.: Chandelier plant, mother of thousands, mother of millions, mission bells, Christmas bells, and friendly neighbour. The latter probably will not be endorsed by the cattle farmers in Australia.
Literature:
Boteler, Thomas (1835). Narrative of a voyage of discovery to Africa and Arabia, Richard Bentley, London.
Ecklon & Zeyher (1837). Kalanchoe delagoensis, Enumeratio Plantarum Africae Australis Extratropicae: 305.
Harvey & Sonder (1862) Flora Capensis 2: 380.
Owen, W.F.W. (1833). Narrative of voyages to explore the shores of Africa, Arabia, and Madagascar, Richard Bentley, London.
Vandenbroeck, F. (1984). De bizarre succulentenwereld van Madagascar, Succulenta 63 (9): 201
Originally published in Succulenta Febr. 2012. Translated from the Dutch by F.N.
Fig. 1 The plants produce miniature plants on the edge of the leaves.
Fig. 2 If the humidity is not too low, little roots develop on the adventitious plantlets even while these are still attached to the mother plant.Fig. 3 Top view of a K. delagoensis with adventitious plantlets.Fig. 4 K. delagoensis has 2 to 9 teeth at the tip of a leaf.
Fig. 5 Anyone who sees K. delagoensis in bloom can hardly imagine that many consider it a troublesome weed.
Fig. 6,7,8 In habitat near Ambovombe. Photos F.N.
(Other photos by Theo Heijnsdijk)

 Fig. 1: The color plate accompanying Jacquin’s 1804 description.
Fig. 1: The color plate accompanying Jacquin’s 1804 description. Fig. 2
Fig. 2 Fig. 3: top view of a flowering plant.
Fig. 3: top view of a flowering plant. Fig. 4
Fig. 4 Fig. 5
Fig. 5 Fig. 6: An almost ripe fruit.
Fig. 6: An almost ripe fruit. Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).IMG_4806 Coby Keizer.
Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).IMG_4806 Coby Keizer.

 Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.
Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85. Fig. 11: Cyathium of Euphorbia clava with female flower.
Fig. 11: Cyathium of Euphorbia clava with female flower.


 Fig. 4. The flower of A. papyracea and a seed pod.
Fig. 4. The flower of A. papyracea and a seed pod.
 Fig. 6. The seeds of A. papyracea are slightly prickly.
Fig. 6. The seeds of A. papyracea are slightly prickly. Fig. 7
Fig. 7





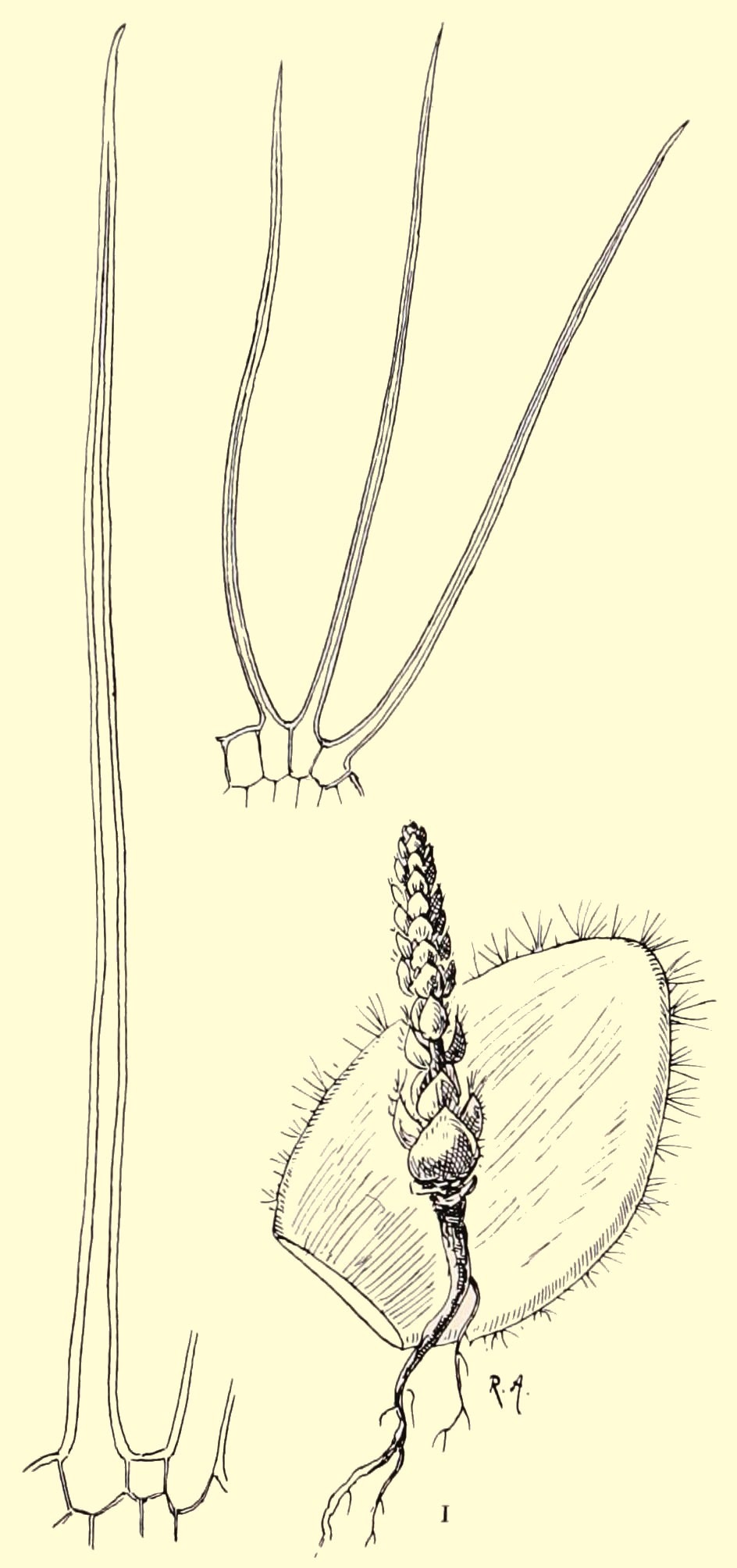
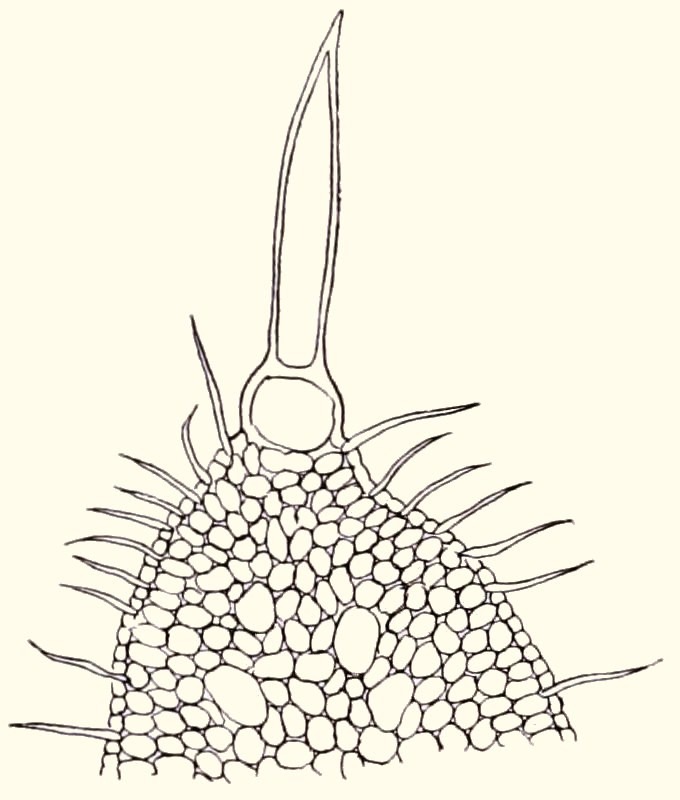






























































 Fig. 1 The plants produce miniature plants on the edge of the leaves.
Fig. 1 The plants produce miniature plants on the edge of the leaves.





