The photos in this post were originally published here as belonging to O. undulosa, but Derek Tribble of London kindly made me aware of them representing in fact O. gymnodiscus. For more information see Derek’s comment.
This is a tuberous, erect or sprawling plant to 35 cm tall, with leathery, oblanceolate to elliptic leaves which are tapering below and somewhat clasping.
The solitary yellow flowerheads are terminal or axillary and disciform ((without ray florets); they appear in July and August.
Its area of distribution is from Cedarberg to Worcester and Little Karoo to as far east as Port Elizabeth on sandy slopes and flats.
Euphorbia clandestina was described in 1804 by Nicolas Joseph von Jacquin (1727 -1817) who was born and raised in the Dutch university town of Leiden. After studying medicine, he moved to Paris, and from there he ended up in Vienna. Apparently, natives of Leiden played an important role in Vienna at that time. The personal physician of Empress (of the Holy Roman Empire) Maria Theresa of Austria, Gerard van Zwieten, was a pupil of the famous Leiden physician Boerhaave. In 1753, on the recommendation of Van Zwieten, the gardener Adriaan Steckhoven, also from Leiden, was appointed director of the gardens of Schönbrunn Palace, also in Vienna. By order of Emperor Francis 1 Stephan, the husband of Maria Theresa, Von Jacquin was sent to Central America and the Caribbean to collect plants, animals, and minerals. The plant collection of the gardens was later described by Von Jacquin in the book “Plantarum rariorum horti caesarei Schoenbrunnensis descriptiones et icones” which appeared in four volumes between 1796 and 1804. Von Jacquin had long since left Schönbrunn. In 1762 he was appointed professor of minerals and mining at the Mining Academy in Schemnitz (now known as Banská Štiavnica). In 1768 he was appointed professor of botany and chemistry and became director of the botanical garden of the University of Vienna.
Outward appearance In the aforementioned four-part series of books, 1270 plants are depicted and described, unfortunately in Latin, which is difficult to understand for most modern readers.
In volume 4 (1804) we find the description with colour image of E. clandestina.
Fig. 1: The color plate accompanying Jacquin’s 1804 description.
The slender stems are 2.5 to 4 cm in diameter and usually up to 60 cm high, but up to 1.2 m has also been reported. The leaves are up to 4 cm long and 2 to 4 mm wide. Older leaves drop off so that only a crown of leaves at the top remains. This gives a good view of the stem, which is covered in spirals of knobbly tubercles with a scar from the dropped-off leaf on the most outward part. See Figure 2.
Fig. 2 This appearance has given the species the name ‘volstruisnek’ (ostrich neck) in South Africa.
Flowering and fruiting The species name clandestina (hidden, inconspicuous) refers to the inflorescence of the species. The greenish inflorescences (cyathia) that appear in spring are situated between the leaves at the top. As shown in figure 3 this produces a beautiful image.
Fig. 3: top view of a flowering plant.
In figure 4 we see a close-up of an individual cyathium in an early stage of flowering. The cyathophylls (bracts) are yellowish-green. They enclose a circle formed by 5 nectar glands. These in turn enclose the flowers proper. In the middle is the female flower (pistil) with 3 incised stigma lobes. On the left, we see a male flower (with the double anthers) that is just about to open and on the right male flowers that have already come out and with their tufts of stamens are somewhat reminiscent of a shaving brush. The species is self-fertile and because male and female flowers are often ripe simultaneously even in one cyathium, pollination is almost guaranteed.
Fig. 4
This results in fruits that are initially green and nearly spherical (fig. 5),
Fig. 5 later turning into an indeterminate brownish shade (fig. 6);Fig. 6: An almost ripe fruit.
it can be seen that the remains of the stamens are still present). Once that stage has been reached, it is high time to take measures to prevent the seeds from being hurled away so that you are later confronted with “clandestine inhabitants” in the pots of other plants.
The natural habitat E. clandestina is quite common in the Western Cape. There, the plants usually grow at altitudes between 60 and 450 meters, on hills among rocks and various kinds of thickets. In Figure 7, a photo by Coby Keizer, we see a specimen (estimated to be 1 meter tall) about 30 km north of the town of Riversdale.
Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).
In Fig. 8, a photo by Bertus Spee taken at De Rust, we see what the sun in South Africa does to the plants: they are beautifully stocky and splendidly coloured.
Fig. 8 Cultivation As far as cultivation is concerned, all sources say that the plant needs a light location and a lot of water and is certainly not very sensitive to cold. In the Western Cape, even light frosts seem to be tolerated. But under our climatic conditions, it is recommended not to allow the temperature to drop below about 5 °C. Sowing gives me varying degrees of success. On June 1, 2014, I sowed three rows of three freshly harvested seeds in a square pot. The three seeds in what I call the front row came up. See Figure 9. Fig: 9: Between the cotyledons of these 3 seedlings the first elongated leaves begin to develop. The other 6 didn’t. On June 10 I sowed 11, again freshly harvested seeds in a 5.5 cm pot. At the end of August, nothing had happened, and I put the pot under the tablet together with other pots with “unfinished business”. On September 20, more than 3 months later, I suddenly saw some greens under the table and it turned out that there were suddenly 7 seedlings present. On the same day, at the foot of the mother plants on the table, I also discovered some newly emerged seedlings. Weather conditions probably play a role here.
Similar species E. clandestina is sometimes confused with E. clava which has stems of a similar form. That species has been known since the year 1700 but was only described around 1781, also by Von Jacquin. Fig. 10 shows the colour plate with the description.
Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.
In the non-flowering state, both species can be distinguished because E. clava has slightly thicker trunks and produces branches from the base, whereas E. clandestina rarely branches, and if it does then not from the base.
But the most obvious is the difference in inflorescence. In E. clandestina, the inflorescences (cyathia) are sessile, that is, without a flower stalk. In E. clava, each cyathium always has a peduncle of 7.5 to 17.5 cm long. In Figure 11 we see a cyathium with a female flower. The flower stalks become woody and remain permanently attached to the plant like a kind of thorns.
Fig. 11: Cyathium of Euphorbia clava with a female flower.
A species that is even more similar to E. clandestina is E. cylindrica. In this species, the stems are somewhat thicker (up to 5 cm) and the cyathia are raised on short stalks.
Literature: Jacquin, N. J (1781 -1786): Icones plantarum rariorum Vol. I: 9, t 85.
Jacquin, N. J. (1804): Plantarum rariorum horti caesarei Schoenbrunnensis. Vol. IV: 43, t. 484 .
Laren, van, A,J. (1932). Succulents, Verkade’s factories N.V., Zaandam, NL.
Originally published in Succulenta 93 (6) 2014. Translated from Dutch by Frans Noltee.
Taxonomy
Avonia used to be a subgenus of Anacampseros, a relatively small genus in the family of purslanes (Portulacaceae).
The species within the genus Anacampseros consist of small to very small shrublets with a more or less thick, sometimes branched stem. The fleshy leaves form a flat rosette or are attached to the stem in a spiral. A typical characteristic are the stipules (lateral outgrowths at the base of the leafstalks). Often these have been reduced to hairs, or grown into parchment-like white scales which in some species can completely cover the stem including the real leaves. In 1994 Gordon Rowley concluded that these species with scale-shaped stipules deserve the status of a separate genus; thus the genus Avonia came into existence. In it, he placed 9 species. There are now 12. The name is probably derived from the Latin ‘Avus’ which means grandfather. It would then refer to the white scales that give the plants an ‘old’ appearance. Rowley placed a few other Anacampseros-like plants in the genus Grahamia. According to his classification, 15 species remain in the genus Anacampseros.
Together, the genera Anacampseros, Avonia, and Grahamia form the branch (tribe) Anacampseroteae within the Portulacaceae.
Incidentally, it is doubtful whether the genus Avonia will hold. It is rather unusual today to base a genus on a similarity in its external appearance. As a rule, the characteristics of the flower and seed and even more so the similarities and differences in DNA play a decisive role.
Flower and fruit
What all representatives of the Anacampseroteae have in common is the construction of flowers and fruit. The flowers, which in all species are only open for a few hours in the late afternoon, always have only 2 sepals, which first completely enclose the flower and often curl up after flowering but remain joined at the tip so that they stay on top of the developing fruit like a cap. This can be seen very well in Avonia quinaria ssp. alstonii (fig.1). At a certain moment the cap falls off and the seeds are released.
Fig. 1
Furthermore, there are usually 5 (sometimes 4) petals , which are white, pink or carmine. The stamens are white with yellow anthers and very variable in number: from 5 in, for example, Anacampseros comptonii to as much as 80 in A. quinaria (fig. 2). The white pistil has 3 lobes. The fruit consists of a kind of veined basket that often has a narrow opening at the top, so that the seeds come out in dribs and drabs as if with a salt-sprinkler as the flower stem bends to the ground and is moved up and down by the wind.
fig. 2
A. papyracea was chosen as the type species of the genus because of its most pronounced characteristics. The name papyracea (=paper-like) is particularly appropriate because a stem looks like a little wad of wafer-thin pieces of paper. The name was given by the German botanist and director of the botanical garden in Königsberg, Ernst Heinrich Friedrich Meyer (1791-1858). The description under the taxonomic rules followed a bit later and is by the Austrian botanist Eduard Fenzl. It appeared in 1840 in a large article about the Molluginaceae (which at that time still included the Portulacaceae) in ‘Annalen des Wiener Museums der Naturgeschichte’. Hence the designation ‘E. Meyer ex Fenzl as found in botanical literature.
The white scales undoubtedly protect the green leaves underneath them against the bright sunlight. Rowley observed that under humid conditions the scales deflect somewhat from the stem so that there can be a better exchange of gases (oxygen, water vapor) with the environment. Perhaps water can also be absorbed in liquid form.
Like most Avonias, A. papyracea hails from South Africa (Western and Northern Cape). It is fairly common and grows mainly in plains with white quartz stones. But the plant also grows on dark coloured soil, where it is of course much more obvious. Figure 3, a photograph by Coby Keizer, shows a cluster in Goegap Nature Reserve east of Springbok.
The former director of the Royal Botanic Gardens at Kew, Sir William T. Thiselton–Dyer, pointed out that the white stems bear a certain resemblance to bird droppings. It is not for nothing that the plant is also called ‘gansmis’ (goose dung) in South Africa. Probably the geese have a different menu there than in Europe, because here the droppings of geese are just as green as the grass they eat. Thiselton–Dyer suggested that this is a mimicry plant, which in this way tries to prevent it from being seen as a tasty snack. If that is the case, then it does not seem to be a successful imitation, because another name for the plant is ‘haasieskos’ (Hare food). The same name is also used for A. ustulata and A. filamentosa. Thiselton-Dyer made his remark in 1906 in an article in the British ‘Annals of Botany’. At that time, the flower was still completely unknown. In the same article he mentions that his friend Nicholas Edward Brown, also from Kew, had been growing the plant for many months, but despite daily observation had never seen a flower on it. Yet, suddenly a fruit appeared from between the parchment-like leaves. He concluded that the flower had to be very small and hidden under the scales had fertilized itself. A cleistogamous species, in other words. Later authors also maintain that the flowers do not open. But that’s not true. Sometimes the plant does bloom and the flowers are not small nor do they remain hidden under the scales (Fig. 4).
Thiselton-Dyer also reported in the article mentioned above that the stalk of the ripening fruit continues to grow so that the seeds are scattered at some distance from the mother plant. This can also be clearly seen in Fig. 4.
Fig. 4. The flower of A. papyracea and a seed pod.
However, this is not always the case. In Fig. 5 it can be seen that the 2 fruits hardly appear from between the scales.
Fig. 5. A full seed pod of A. papyracea and an already emptied one. Both only just visible between the scales.
In addition to the usual species which is nowadays referred to as A. papyracea ssp. papyracea, there is also a subspecies: A. papyracea ssp. namaensis. This occurs in the northwestern part of South Africa and in the south of Namibia and is distinguished by the fact that the scales are not smooth-edged but serrated (saw-edged) or toothed and by the seeds that are lumpy. In ssp. papyracea they are more prickly (Fig. 6).
Fig. 6. The seeds of A. papyracea are slightly prickly.
Seed collection is extremely simple. Holding an empty tube or something similar under a ripe fruit and a tap against the fruit are enough. Immediate sowing gives excellent results. At a temperature of 25 to 30 °C, the seedlings emerge en masse after a week. Only a short time later, the seedlings proceed to develop the papery scales (Fig. 7).
Fig. 7
A year later they have become miniatures of the mature plants ( see fig 8).
Fig. 8
Cultivation
I grow the plants in standard cactus soil and I also treat them like cacti. This means that they receive water regularly in summer and that I keep them dry in winter. I try to keep the temperature in my greenhouse around 8 °C in winter, but it sometimes drops to just above freezing point. According to the literature, even temperatures of 5 degrees below zero are survived.
Apparently, A .papyracea, like A. ustulata, was also used in South Africa as a kind of yeast for baking bread and making beer. To this end, roots and stems are dried and ground. It is suspected that the plant also contains psychoactive substances, just as, by the way, A. quinaria. The brew must have been a kind of ‘spacebeer’.
Literature
Browne, P. (1756). The Civil and Natural History of Jamaica. In Three Parts [3]: 234
Fenzl, E. (1840). Monographie der Mollugeneen, Annalen des Wiener Museums der Naturgeschichte, [2]: 295
Marloth, R (1917). Dictionary of The Common Names of Plants, The Specialty Press of South Africa, Ltd. Cape Town
Rowley, G. (1994). Anacampseros and allied genera- A reassessment, Bradleya 12: 105-112
Sims, J. (1811) Anacampseros filamentosa, Curtis’s Botanical Magazine 33: plate 1367
Thiselton-Dyer, W.T.(1906). Morphological Notes, Annals of Botany 20 (2) blz 123-127
Originally published in Succulenta 93: 2014. Translated from Dutch by FN.
Plants of var. obtusifolia have many stiff woody branches up to 30 cm (usually rather smaller); the leaves are yellowish green and up to 1 (sometimes 2) mm long and wide.. They flower in August-November.
Occurring from southwestern Namibia just north of the Orange River to the Cederberg in South Africa, usually in crevices or shallow soil on rocky outcrops.
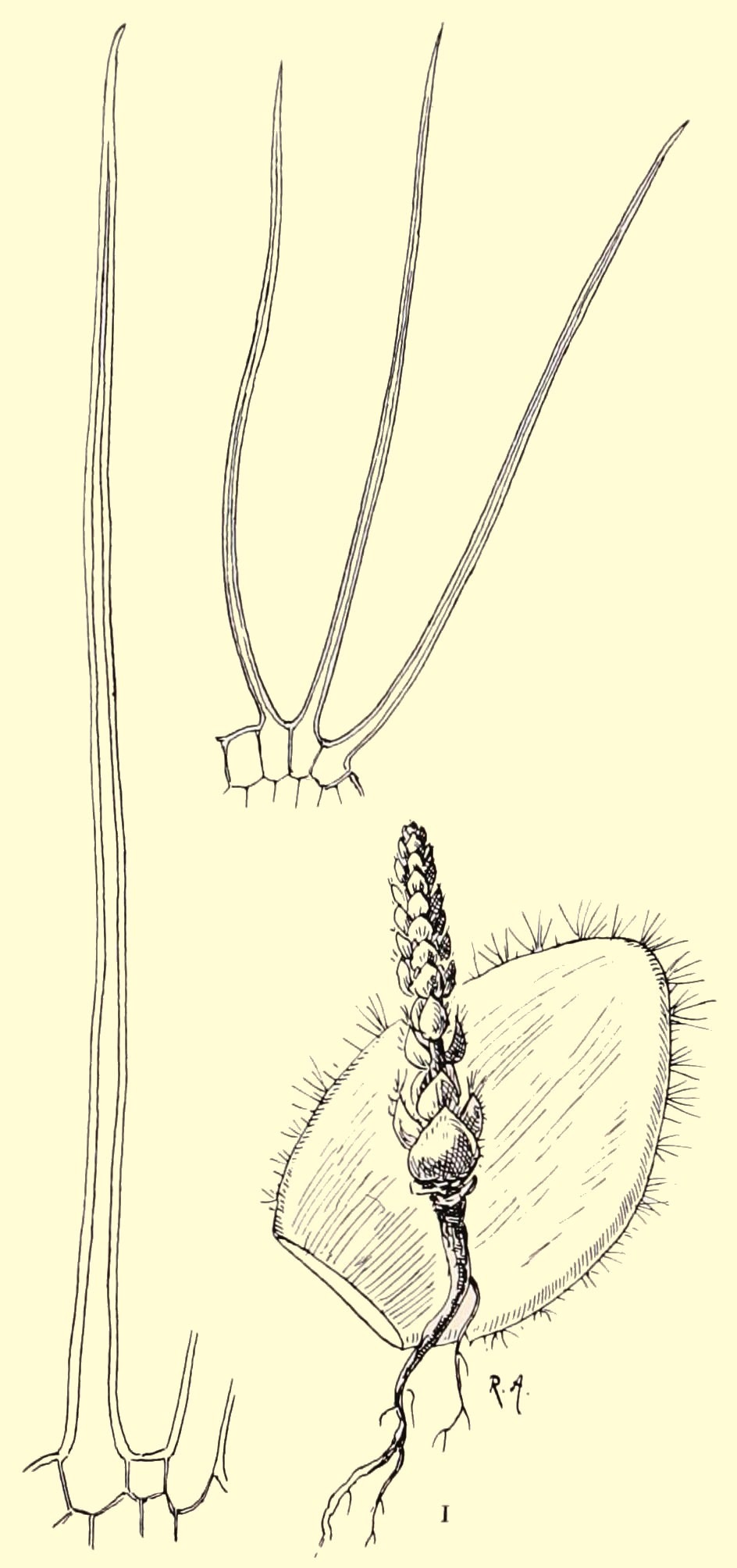
C. barbata is among the 28 new South African Crassula species described by Carl Thunberg in 1778. In nature it occurs on rocky slopes in quite a large area including, among others, the Great Karoo and the Little Karoo. Characteristic for the species are the up to 5 millimeter long hairs on the leaf margins which give the impression of entangled eyelashes (fig. 1) or a beard – as the the botanical name implies (barbata = with a beard). At the tips of the leaves the hairs are often gathered in little tufts (fig. 2). Apart from this, the leaves are completely hairless. In nature, the plantlets often look like little fluffy globes. See fig. 3, a picture made by Coby Keizer in November 2008 at a spot about 40 kms west northwest of Matjiesfontein where a lot of interesting little succulents grow together.
Fig. 1Fig. 2
Fig. 3
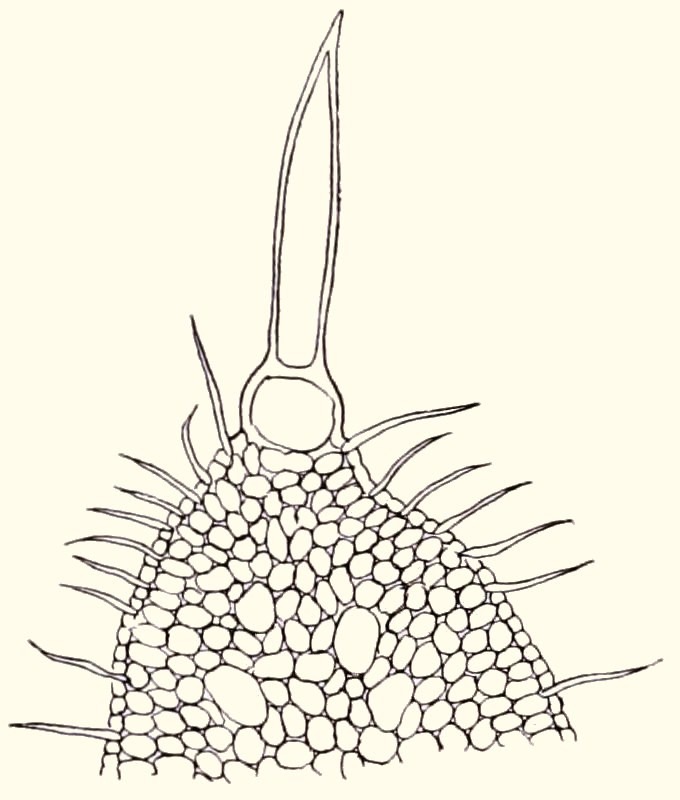
In the guest column on CRASSULA PYRAMIDALIS *, I mentioned the experiments that Rudolph Marloth carried out early last century to assess water absorption by Crassulas through the leaves and hairs. A plant of C. barbata gained 5.2% in weight in one night by absorbing dew. Marloth also looked at what happens when you apply a small drop of water to the end of a hair on a wilting leaf. Before long, the hair swells and stands upright and gradually the leaf regains its turgor. In Crassula tomentosa (fig. 4), with its row of thick hairs, this works even better. Marloth took a leaf from it and dipped the hairy edge in the water. After 12 hours, the weight had increased from 14.45 to 16.05 gram, an increase of no less than 11%.
Fig. 4 Marloth’s article was illustrated with several beautiful drawings in which you can also see that the hairs have developed from epidermis cells (fig. 5 and fig. 6).
fig. 5fig. 6
C. barbata is a slow grower. It takes a few years until the heart of the rosette opens and the beginning of the flower stem becomes visible (in the Netherlands usually at the end of November). At this stage, the flower stem looks like a C. pyramidalis plantlet (Fig. 7).
Fig. 7 Fig. 7 The stem grows rapidly (fig. 8 and fig. 9) and after a few weeks the maximum height (up to 30 cm) is reached.
f
Fig. 8
Fig. 9 At the bottom there are still leaves with lashes. The flowers are arranged in groups of about 7 and several of those groups stand in a crown around the stem. The petals are usually beautifully white, and this contrasts nicely with the yellow anthers (fig. 10).
Fig. 10
Pink shades also occur . Fig. 11
The flowers emit a pleasant, sweet smell.
There is only one known variant of C. barbata: ssp. broomii, also known as an independent species: C. broomii. It grows in the vicinity of Victoria West (North Cape) in the mountains and is distinguished from the type plant mainly because the hairs are considerably shorter, up to only 1 mm. For plantlovers this makes it not so interesting, because it is precisely the hairs that make the plant so attractive. We therefore hardly ever encounter this variety in collections.
There are also hybrids of C. barbata, but they are not common in culture. In an article in Succulenta (1980), B.K. Boom mentions a cross with C. orbicularis cv. ʽRosula’ and also shows a photo. Rowley mentions a cv. “Roger Jones” as being “C. barbata x ? orbicularis”, but does not give an image.
Gordon Rowley, in his book ‘Crassula’, considers C. barbata a joy to see in its natural habitat but usually a disappointment in cultivation. Instead of remaining compact, the plants grow into elongated loose rosettes and after flowering the plant dies. But he also made this kind of statement about C. columnaris and C. pyramidalis. My experience is that it is not as bad, as long as you are economical with water. Watch out for total dehydration in summer and give the plant the lightest spot you can find in winter.
Also, the plant does not always die after flowering. For example, I had a plant that bloomed in the winter of 2011-2012. As the rosette died off, 5 new shoots formed at the base of this plant. At the moment, at the beginning of December 2013, a flowering stem rises above the rosette in all 5 of them. So, you can enjoy such a plant for years.
Literature: Boom, B.K. (1980). De Crassulas van onze collecties 6 and 7, Succulenta 59 (1): 120. Laren, van, A,J. (1932). Vetplanten, Verkade fabrieken N.V., Zaandam. Marloth, R. (1908). Das Kapland, Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer “VaIdivia” 1898-1899, bd.2, t.3. Noltee, F. (2008). Succulenten van de Karoo, een bezoek aan Perdekraal, Succulenta 87 (4): 170 and Succulenta 87 (5): 215. Rowley, G. (2003). Crassula, Cactus & Co. Thunberg C.P. (1778). Nova acta physico-medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum 6: 328-341.
Originally published in Succulenta 93 (4) 2014. Translated from Dutch by Frans Noltee.
Small plants of this species could be mistaken for C. elegans ssp. elegans, but that taxon* usually has many branches, with more open tips.
The species also resembles C. plegmatoides, which however has coarse (usually rounded) papillae on the leaves.
Usually with erect, up to 15 cm tall stems, often with a few branches.
The leaves are tightly packed, broadly egg-shaped, 0.5-0.9 x 0.7-1.3 cm, with bluntly acute, often mucronate** tips, concave above and convex below, velvety, green to yellowish green, but often tinged red towards the tip; they are closely clasping around the stem, forming a four-angled column 0.7-2 cm in diameter.
The tubular, cream to pale yellow flowers appear from Februari to April.
Found in Namaqualand between Grootmis and Lekkersing and north to the Buchu Mts. in Namibia, usually growing in crevices in quartzite outcrops on the southwest facing slopes of low hills.
* taxon: a term which can be used to refer to any taxonomic group, depending on the context.
**mucronate: abruptly ending with a small, three sided tip, which is sometimes extended into a bristle or spine.
Over time these plants form tight clumps often half-hidden in the ground, 3-4 cm tall and 5-10 cm across.
The leaves are short, united for up to two thirds and hardly keeled; the surface is obscurely warty (= verrucosa) with conspicuous dark spots.
In late winter-early spring ( August-September) they produce small yellow flowers (up to 2.5 cm across) on short stalks.
The species is endemic to the northwest corner of the Richtersveld, an area with less than 100 mm rainfall per year (in winter). Usually growing in deep sand-filled cracks of shale or dolomite.
The year 1778 is an important one for Crassula lovers. In that year, an article appeared in the Swedish periodical with the wonderful name ‘Nova Acta Physico-Medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosum’ in which physician-botanist Carl Peter Thunberg (also from Sweden) briefly described no less than 28 new South African species of this genus. Most of these have become well-known among fanciers. Just think of Crassula alpestris, C. barbata, C. capitella, C. columnaris, C. corallina, C. hemisphaerica, C. tecta, C. tomentosa, and the subject of this article: C. pyramidalis.
At the back of Thunberg’s book is a plate depicting 2 of the new species, C. alpestris and C. pyramidalis (see fig. 1).
fig. 1
In later years, of course, many more images of C. pyramidalis appeared. Particularly beautiful is the coloured plate published in 1899 in Curtis’s Botanical Magazine (fig. 2). Here we see a thicker shape than Thunberg’s plant has.
fig. 2
C. pyramidalis is closely related to C. columnaris (discussed earlier in this series) and to C. barklyi. All three of them are highly succulent species whose leaves are so tightly packed together that the stem is totally invisible. In C. pyramidalis, the leaves are grouped very regularly. At the top of a stem, this results in a particularly beautiful view (fig. 3).
fig. 3
In nature, the plants can take on a beautiful purple-red color (See figure 4). Coby Keizer took this picture in November 2008 about 60 km north of Matjiesfontein (Western Cape) along the road to Sutherland.
fig. 4
Not visible in photos are the hairs in the leaf axils. They allow the plant to absorb water. In the Dutch picture album of the Verkade Company this is described by the author, A. J. van Laren, as follows: ‟Remarkable in some of these Crassulas is the ability to absorb water by means of hairs on the leaves, such as the marginal hairs of C. barbata; in C. columnaris and C. pyramidalis there are such water-absorbing hairs at the base of the leaves, thus at the trunk. The effectiveness of these hairs is rather significant! To give an example, a leaf of C. barbata can absorb more water in one night, as a result of dew, than it loses in a week by transpiration. The placement of the leaves in C. pyramidalis is such that water flowing down the plant penetrates into the spaces between the leaves, from where it is channeled to the base and absorbed by the hairs present there. When drought sets in, the leaf margins are so close together that the water-absorbing hairs at the base of the leaves are completely closed off from the outside air and therefore cannot lose water.
I suspect that van Laren derived the above information from the book which the German chemist/pharmacist/botanist Rudolph Marloth (1855 -1931) wrote under the title ‘Das Kapland, insonderheit das Reich der Kapflora, das Waldgebiet und die Karroo’. This book with over 550 pages in 1908 as part 3, volume 2 of a whole series under the title ‘Wissenschaftliche Ergebnisse der Deutschen Tiefsee-expedition auf dem Dampfer “VaIdivia” 1898-1899’. The Valdivia was a steamer commissioned by the German government to undertake a deep-sea expedition in the Atlantic and Indian Oceans from August 1898 to May 1899. The report under the title mentioned above included 24 volumes, the first of which appeared in 1902 and the last in 1940. Marloth, who lived and worked in South Africa from the age of 28 until his death, did not participate in the expedition but became involved by the botanist Wilhelm Schimper who disembarked on the return trip to Cape Town to continue with botanical fieldwork. The concept of deep-sea was apparently understood very broadly. When Schimper died in 1901, Marloth was asked to complete the volume about South Africa.
In his book, Marloth reports on experiments with various Crassula species. He took 2 specimens of C. pyramidalis of which he wrapped the roots in tin foil so that they were closed off from the open air. He put the plants outside for a night. One covered with a piece of cardboard, the other one unprotected. The covered specimen lost a bit of weight (0.6%), whereas the unprotected specimen exposed to the dew increased in weight by 5.1%. Marloth concluded that the plant can absorb 5.7% of its body weight from dew. Of course, that does not prove at all that this is caused by those leaf axil hairs, but it is still a nice fact. According to Marloth, C. columnaris also has such hairs in the leaf axils. In his experiment, this species was found to experience 6.3% weight gain due to the dew. For C. barbata, the increase was 5.2%.
In addition to the position of the leaves, C. pyramidalis, C. columnaris and C. barklyi also correspond in the terminal, almost spherical inflorescence without visible flower stem (fig. 5) so that it seems as if a globule of flowers is crammed into the plant from above. The visual disadvantage is that the dried plug remains in place for a long time after flowering.
fig. 5
C. pyramidalis tends to allow all stem ends to bloom at once. A beautiful sight, but also the harbinger of a rapidly approaching death. This happened to me, for example, with the plant in fig. 6 that I bought at an open day at the Ubink nursery.
fig. 6 The photo was taken 3 months after the purchase. The stems were nicely thickened and coloured slightly red by the sun. At the end of February 2012 I saw that the formation of buds had started (fig. 7).
fig. 7
On April 1, the first flowers were open and on April 9, all stems were in bloom (fig. 8). The flowering lasts for many weeks, but the flowers gradually turn brown, after which the same thing happens to the stems themselves and then the plant is done for.
fig. 8
Thunberg found C. pyramidalis at the thermal springs on the Olifants River (Cape Province). Later, the species was found in many other places in the Cape Province (Little Karoo, Great Karoo, Namaqualand). The species is widespread in other words.. We can also come across the plant in literature under one of the following names: Crassula quadrangula, Tetraphyle pyramidalis, Tetraphyle quadrangula, Tetraphyle pyramidalis var. quadrangula, Purgosea pyramidalis, Crassula archeri and Crassula cylindrica.
In his 1778 article Thunberg wrote that C. pyramidalis resembles C. muscosa, but is distinguished by the thicker stems and the terminal inflorescence. C. muscosa is the valid name for the plant that probably most of us know as C. lycopodioides, the well-known shoelace plant. In this species, the inconspicuous and (to my mind) unpleasantly smelling flowers appear everywhere along the stems between the tiny leaves (fig. 9). Muscosa, by the way, means: moss-like, referring to the collection of thin stems. Lycopodioides means: resembling wolf’s claw (Lycopodium).
fig. 9
Over the years, a number of hybrids of C. pyramidalis have become available. Without a doubt the best known is ‘Buddha’s Temple’, a hybrid with C.perfoliata var. falcata. A photo of this plant in side view was already shown in the article in this series about the latter. In fig. 10 this plant is shown in top view.
fig. 10
We also know ‘Giant’s Watch Chain’ (C. muscosa x pyramidalis), ‘Starburst’ (C. ausensis x pyramidalis), and the multihybrides ‘Star Child’ (C. deceptor x ‘Starbust’) and ‘Tinkerbell’ (C. ausensis x ‘Springtime’) x pyramidalis) in which ‘Sprintime’ in its turn is a cross between C. perfoliata var. falcata with (probably) C. rupestris. A hybrid of no less than 4 species. It will probably make your head spin in the meantime.
A hybrid that also deserves a place here is C. ‘Extagnol’ which we also come across under the name C. ‘Estagnol’. This plant stands out because of the spiraling leaf position. In close-up (fig. 11) it reminds one somewhat of Aloe polyphylla .
fig.11
But remember that the diameter of this plant is only 10 mm. Its origin is not clear, but in the literature it is stated that it is a cross between C. pyramidalis and C. perfoliata var. perfoliata. I think the leaf shape is more indicative of C. alpestris, but that is only a random guess of course.
In cultivating C. pyramidalis and all these hybrids, we must bear in mind the highly succulent character of the plants. That is to say: a mineral soil, a sunny location, careful with water and little to no fertilizer.
Literature
Heijnsdijk, T. (2013). Crassula perfoliata var. falcata, Succulenta 92 (1): 3.
Heijnsdijk, T. (2013). Crassula barklyi, Succulenta 92 (4): 178.
Hooker, J. D. (1899). Curtis’s Botanical Magazine 125, tab 7665.
Laren, van, A,J. (1932). Succulents, Verkade’s factories N.V., Zaandam.
Marloth, R. (1908). Das Kapland, Wissenschaftliche Ergebnisse der Deutschen Tiefsee-expedition auf dem Dampfer “VaIdivia” 1898-1899, bd.2, t.3.
Thunberg, C.P. (1778). Nova acta physico-medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum, t6: 328-341
Images:
Fig. 1 The first published image of C. pyramidalis (Thunberg, 1778)
Fig. 2 A part of the image in Curtis’s botanical from 1899
Fig. 3 A young side shoot
Fig. 4 C. pyramidalis approx. 60 km north of Matjiesfontein along the road to Sutherland. (photo Coby Keizer)
Fig. 5 The almost spherical inflorescence of C. pyramidalis.
Fig. 6 C. pyramidalis in a 5 cm pot
Fig. 7 Button formation at the plant of fig. 5 C. pyramidalis
Fig. 8 The same plant: all shoots bloom at the same time
Fig. 9 Crassula muscosa (lycopodioides) in bloom
Fig. 10 Crassula ‘Buddha’s Temple’ (C. perfoliata var. falcata x pyramidalis)
Fig.11 Crassula ‘Extagnol’ (possibly a hybrid of C. pyramidalis and C. perfoliata var.perfoliata)
Originally published in Succulenta 93 (3) 2014. Translated from Dutch by Frans Noltee.
This species is a common sight on soft saline soil and low outcrops in the Richtersveld, from north of Lekkersing to the Augrabiesberg, at an altitude of 200-300 m. This is a winter rainfall area, which receives less than 100 mm rain per year.
It develops large, dense clumps 6-10 cm tall and up to 50 cm in diameter.
The more or less erect leaves are pale greyish white to bluish green, broad and thick (1-2.5 cm) and up to 4.5 cm long.
The flowers are 5-6 cm in diameter and range from cream and yellow to orange-red, sometimes with a purplish tinge. They appear in winter and spring (August-September).
This attractive species is widespread in the summer rainfall area of South Africa from the North-eastern Karoo into the Free State and Bushmanland and is also found in southern Namibia. It occurs in flats, in shale or coarser sand or gravel and easily establishes itself along roadsides and in other open spaces.
The plants are rather variable: compact to freely branched with creeping branches, forming dense mats or growing in tufts.
The leaves are more or less trigonous, 1-2.5 cm long, 0.4-0.5 cm wide and thick; they are covered with prominent whitish warts.
Flowers appear from August to December; they have a very short stalk and are about 1.5 cm in diameter. The petals are golden yellow but reddened at the tip on the back surface. They open from late afternoon to early evening.
NB. The picture labelled C. musculinum in Robin Frandsen’s “Succulents of Southern Africa” does not show this species but rather a Stomatium.






 Fig. 1: The color plate accompanying Jacquin’s 1804 description.
Fig. 1: The color plate accompanying Jacquin’s 1804 description. Fig. 2
Fig. 2



 Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).
Fig. 7: A strapping Euphorbia clandestina in the Western Cape (South Africa). (photo Coby Keizer).

 Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.
Fig. 10: The colour plate accompanying Von Jacquin’s description of Euphorbia clava. Table 85.


 fig. 2
fig. 2


 Fig. 6. The seeds of A. papyracea are slightly prickly.
Fig. 6. The seeds of A. papyracea are slightly prickly. Fig. 7
Fig. 7







 Fig. 3
Fig. 3