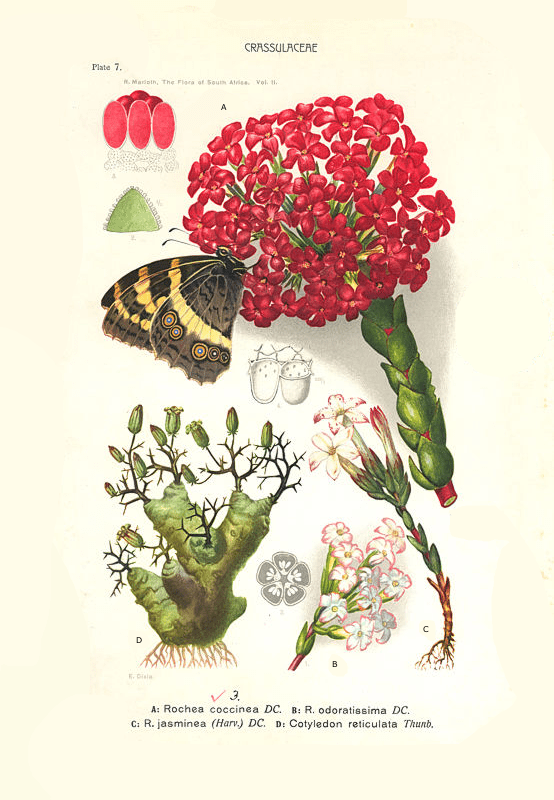This species was introduced in the Netherlands as far back as 1700 and is the ancestor of the peltate-leaved cultivars. It is quite variable in the size, texture and colouring of the leaves and makes an excellent garden plant which grows easily from cuttings. In nature it is widespread from Wellington and Worcester to Kwazulu-Natal.
It is a climbing plant with soft slender stems up to 4 m long, which trail into and through other bushes.
The leaves are 2-7 cm in diameter with peltate (shield-like) blades, which are round to 5- to 7-angled or -lobed; usually glabrous (with a smooth surface) and sometimes decorated with zonal markings.
Flowers are 4-5 cm in diameter and arranged with 2-9 per inflorescence. They have 5 purple to light pink or even whitish petals; the upper two have darker markings and are much wider than the lower three.
Flowering time is spring and summer (mainly Sept. – Jan.)
At first glance it is often easy to mix up this species with E. schoenlandii (see link Euphorbia schoenlandii).
This applies especially to young plants. In mature specimens the easiest way to tell them apart is by looking at the peduncles (main flower stalks): in E. schoenlandii these become sharp, woody spines, whereas in E. fasciculata this is not the case.
The plants are unbranched, glabrous, up to 30 cm tall with a 3-8 cm thick, cylindrical stem. This stem is covered with spirally arranged, large hexagonal tubercles, each with a characteristic triangular central depression.
The leaves are long and narrow (1-2.5 x 0.2-0.4 cm) and short-lived.
Plants of E. fasciculata are monoecious, which means that the flowers are either male or female, but both occur on the same plant.
The cyathia are gathered around the tip of the stem, several on each peduncle, 0.6-0.7 cm in diameter and with 4-5 green glands. They appear from June to October.
Known from less than 10 localities from the Vanrhynsdorp area southwards to Clanwilliam, the species occurs in loam, sand or quartz gravel on flats or gently sloping hills.
C. columnaris was described as early as 1778 by the Swedish physician and botanist Carl Peter Thunberg. He has been called ‘the father of South African botany’ and ‘the Japanese Linnaeus’. From 1772 to March 1775 he was in South Africa and it is certain that in September 1772 he collected plants in the Little Karoo, one of the areas where this species occurs. From 1775 to 1778 he was a medical doctor in Deshima, the Dutch settlement in Japan. On his way to Amsterdam, he spent another 2 weeks in South Africa in 1778. It is plausible that Thunberg collected the plant himself or at least saw it in its natural habitat and therefore it surprises me that he gave it the name columnaris (= columnar).
In nature, C. columnaris is almost spherical, flat rather than elongated. It has been noted several times in literature that the species better could have been named C. globosa (= spherical). Perhaps Thunberg continued to cultivate the plants that he took with him for a while, resulting in them losing their natural appearance. This is a problem that these types of plants tend to suffer from in cultivation.
English-speaking people often use the name ‘khaki button’ which, in addition to the shape which the plant assumes in the dry season, also refers to the distinctive color that the plants adopt (see fig. 1. Photo Winfried Bruenken).
Fig 1
The Afrikaans name ‘koesnaatjie’ for this plant also refers to the flattened convex appearance. A naatjie, also called naartjie, nartjie or nartje is a small citrus fruit that is halfway between a mandarin and a tangerine. Koes means in hiding, withdrawn. A mandarin in hiding more or less, probably referring to the way in which the plant partially withdraws into the ground in the dry season (I assume that the plant does not taste like a mandarin, even if it is edible). On the internet I found the following description of the koesnaatjie: “A koesnaatjie is a succulent plant found in the dry western and southwestern regions of the Cape. The plant has thick, fleshy, oval-shaped leaves which are fused at the base and fold tightly over each other like roof-tiles, so that the plant has a compact appearance, and it is edible. Only once does it develop an inflorescence with densely congregated cream to yellow flowers after which the plant sets seed, withers and dies”.
In the 20’s and 30’s of last century, the Dutch monthly magazine ‘Succulenta’ regularly wrote extensively about this plant and the differences between nature and cultivation were always pointed out in detail. For example, in the November 1935 issue I read:
Miss W. v. A. in B. writes: “Can I take cuttings of an etiolated Crassula columnaris? They are so ugly with that brown piece of stem. What causes these plants to grow so spindly here?”
And the editors’ answer:
“The proper cultivation of highly succulent Crassula species in our climate is not easy. Many of these plants originate from the Karroo highlands. The average rainfall there is 4.47 inch ( ± 114 mm). In our country, an average of 700-800 mm rainwater falls per year. The rain on the Karroo plain falls mainly in the four winter months, sometimes also in November and December. These two months of summer rain have little influence on growth, but are important for replenishing the water reservoirs of the succulents.
Few deserts have a more desolate appearance in the dry period than the Karroo. There are no green plants to be found, the gullies are dry and the isolated “Koppies” (small mountain peaks) which are scattered here and there in the Central Karroo, seem to reflect the heat like a mirror. During the night a cool S.E. breeze prevails. The imaginary line which divides the districts into regions with summer or winter rains, runs through the Central Karroo near Beaufort West. In some winter nights the temperature drops below freezing point, and the mountain peaks are sometimes covered with snow, which, however, rarely falls on the plateau itself. The average daily temperature difference is large, i.e. 30° F (= 17 °C). Due to the intensely dry air, the summer heat is not very oppressive. In the Northern Karroo the average rainfall per year is even less, in the Western part 2 inches, so 51 mm per year. The average summer temperature in the shade is 110° F (= 43 °C). This gives us an idea of the conditions under which the highly succulent Crassulas grow in their homeland and makes us understand that it is therefore not easy for these plants to maintain their beautiful, stocky appearance here. Crassula columnaris occurs near Whitehill, the first train stop past Matjesfontein. Jacobsen mentions Namaqualand as habitat. The annual rainfall there is about 6 inches = ± 150 mm. Crassula columnaris allows you to take cuttings (the best time for this is August because the plant starts to grow in this month. After rooting, be very, very economical with water. The entire structure of the plant indicates moisture absorption from the air. In summer provide as much sun and fresh air as possible; the plants may turn reddish brown.”
It is remarkable that between 1923 and 1942, no less 5 times was written extensively about C columnaris in ‘Succulenta’ and after that, which is now over 70 years, nothing at all. The name is only mentioned in passing once in a while.
As can be seen from the above, C. columnaris belongs to the group of winter growers, which have their resting period in summer and should receive minimal water then. In many books a winter temperature of at least 15 °C is recommended, but in my experience that is not good at all. Such a high temperature combined with the application of water promotes growth and with the short days and the low light intensity in winter, this means an elongated, unnatural shape. I have never noticed that the temperature in my greenhouse – which I try to keep at least 8 °C in the winter (with varying success) – would harm the well-being or flowering of such plants.
Other members of the group of winter growers are C. alstonii, barbata, deceptor and pyramidalis. They are best grown in sandy soil with a small proportion of humus.
C. columnaris is monocarp, which means that the plant dies after flowering. This prompted Gordon Rowley in his book ‘Crassula’ to describe cultivating C. columnaris as “a passing pleasure”. Beautiful plants of the species are rare in our collections. Seedlings become elongated and imported plants immediately bloom and then die. To keep the plants somewhat presentable, Rowley recommends a starvation diet with plenty of sun and fresh air.
Although the first signs of flowering also announce the impending death of the plant, it is still fascinating to see how the inflorescence develops. It starts with the closed rosette bursting open at the top, as it were, to provide room for a compact arrangement of flower buds. See Fig. 2.
Fig. 2
This is a plant I bought as a young seedling with the name “C. columnaris Lemoenshoek” at the Special Plant Market in Nijmegen. Lemoenshoek is a farm about 40 km east of the village of Barrydale in the Little Karoo. The photo was taken on December 3, 2012. The diameter of the plant in the photo is 4 cm and its height is about 3 cm. In those 3 cm, 7 pairs of leaves are pressed together. Once in full bloom, the plant body is completely hidden from view by the numerous white flowers. See the photo of Fig. 3, which was taken on January 19, 2013. The name ‘shaving brush crassula’ used in English-speaking countries needs no further explanation.
Fig. 3
Fig 4 C. columnaris ssp. prolifera in the Knersvlakte (photo by Sofia Etzold, with thanks to “Photo Guide to Plants of Southern Africa”). In Afrikaans, this variety is called ‘sentkannetjie’. ‘Sent’ in this expression is derived from the English ‘scent’ and kannetjie stands for bottle. This does justice to both the plant’s shape and the flowers’ sweet smell.
Hybrids of C. columnaris are uncommon, but H. R. Toelken in the Flora of Southern Africa mentions a putative natural hybrid C. alpestris ssp. massonii x C. columnaris ssp. columnaris and gives the following information:
“The plants superficially resemble C. columnaris subsp. columnaris with its clasping leaves producing an erect column c. 15 mm in diameter. However, the plants are longer than broad and have terminal and axillary inflorescences and the leaves are bluntly acute as in C. alpestris ssp. massonii. The putative parents have been recorded from the area”.
Finally: C. columnaris should not be confused with C. columella, which has many similarities with C. columnaris in name and appearance. This plant forms thin, branching, square columns of up to 1.5 cm thick and 5 cm high (columella means ‘small column’), consisting of tightly packed (tile-like) leaves. See below.
Fig. 5
Crassula columella also flowers terminally but does not die afterwards. Next to the faded head (right), one or two new shoots arise. It also flowers terminally, with small, inconspicuously colored flowers on a stalk. Unlike C. columnaris, the stem does not die after flowering but forms 1 or more side shoots directly next to the flower stem, so that a low bushy plant is developed. For us, plant fanciers, this has the great advantage that the plant not only becomes more beautiful after flowering, but also provides a lot of easily rooting cuttings. This species is also hardly prone to the etiolation that C. columnaris so easily suffers from.
Literature:
Laren, van, A,J. (1931). Succulents, Verkade’s factories N.V., Zaandam.
Rowley, C. (2003): Crassula, Cactus & Co.
Swüste, F (1935). Vragenrubriek, Succulenta 17 (11) p. 175.
Thunberg C.P. (1778). Nova acta physico-medica Academiae Caesareae Leopoldino-Carolinae Naturae Curiosorum.
Photo Guide to Plants of Southern Africa, www.southernafricanplants.net
First published in Succulenta 92:5, October 2013. Translated from Dutch by FN.
For more habitat pictures and other info follow this link: Crassula columnaris (part 1 of 3)
Plants of this species are always covered with spines, which makes them easy to recognise. Another helpful characteristic (when you have a living plant in front of you!) is that the plants will smell of fish when damaged.
Mature plants develop into erect shrubs to 25 cm tall, with dark brown to grey internodes and trigonous , 1-1.5 cm long leaves.
Each inflorescence has up to 18 brittle spines on the outside and in July-August may contain1-7 flowers 1-2.2 cm in diameter.
The species is locally common in open Karoo vegetation on stony shale soil from the Cederberg Mountains to Worcester, Matjiesfontein and Montagu.
One could describe this species as a “Big brother” of the earlier discussed Ruschia grisea. It is a robust shrub to 70 cm tall with ascending branches. The leaf-pairs are fused to each other below, with the free parts usually shorter than the basal leaf sheath; the keel is decorated with a single indistinct tooth.
In late spring/early summer (Oct.-Jan.) the plants are blooming with white flowers 22 mm in diam.
The species is to be found on shale and gravel plains of the southern Great Karoo from Matjiesfontein to Prince Albert.
At present, the genus Chasmatophyllum contains nine species, but a proper taxonomic study would probably reduce this number. The genus name is derived from two Greek words: chasma (open mouth) and phyllum (leaf).
C. stanleyi is a low shrub 7-9 cm tall with a stout woody rootstock and ascending branches.
The leaves are trigonous, 10-13 mm long, 3 mm wide and up to 4 mm thick, covered with little warts; the keel usually has a prominent recurved tooth below the tip and the margins sometimes have a tooth on each side.
In September -October the flowers appear; they are yellow (reddish at the tips) and 20-24 mm in diameter.
Occurring on stony to shaley slopes and flats from Laingsburg into the Karoo and the Eastern Cape.
Guest column by Theo Heijnsdijk (Translation and habitat pictures by FN)
History In 1812 Adrian Haworth in his “Synopsis plantarum succulentarum” described a new genus in the family Euphorbiaceae: Dactylanthes, translated by himself as Finger-flower. He distinguished four species in the genus: D. patula, D. anacantha, D. tuberculata and D. hamata, characterized by the finger-like lobes of the inflorescences.
In 1823 he added a fifth species: D. globosa. In the description, he reported that he had seen the species in bloom in Kew in October of that year.
Fig. 1
In 1826, Curtis’s Botanical Magazine published a coloured plate (plate 2624, see figure 1) with a detailed description by John Sims. This time under the name Euphorbia globosa, which is now generally recognized. The genus Dactylanthes has thus merged into the genus Euphorbia. The species D. anacantha and patula are seen as synonyms of Euphorbia tridentata by current botanists. For the other three, the specific name has remained the same.
Habitat This distinctive plant seems to have been widely distributed in the Eastern Cape area of South Africa*, but nowadays the species is much rarer. It occurs in the area between Port Elizabeth and Uitenhage where it grows on low stony hills up to 20 km from the coast.
On the Red List of South African plants, the species has the status ‘endangered’. According to that report, it is known from less than 5 localities. The plant is listed in Appendix 2 of CITES.
* In the famous work “The succulent Euphorbieae” (1941), by White, Dyer and Sloane, we find (among many other things) the following observation on this topic: “E. globosa is abundant in the karoid scrub between Uitenhage and Port Elizabeth, sometimes forming uniform mats of the dwarf branch-tips in such masses that it is difficult for the passer-by not to tread on them.”
Description
It forms compact clumps up to 8 cm high and to 50 cm in diameter, consisting of globular** stem segments that seem to be connected in a rather random way.
** Hence the name globosa (= spherical).
The young shoots bear small scaly leaves which later disappear. The oldest segments are grey to almost white.
African names for the species are eierpol, knopmelkbol and langbeentjie.
A mature segment develops a thin stem that bears the inflorescence, which is supported by 3 bracts. When the inflorescence or fruit has dropped, the flowering stem often divides again into 1, 2, or 3 new stems which like the older ones may become about 8 cm long.
Fig. 2
Fig. 2 shows the scar of the dropped inflorescence, and it is also visible how 2 new stems develop alongside. At the bottom, the third one just starts. A trichotomous division I would say. In this way, a cascade of stems is created. The dried flower stalks remain on the plant. White, Dyer and Sloane have the following comment on the species:“Much has been witten about E. globosa since 1826, yet its “singularly whimsical growing habit” remains as fresh a source of surprise to-day as ever. In its native surroundings the globose branches form thick mats, closely pressed to the ground. These branches are gradually drawn down almost perpendicularly underground, new branches replacing them at the surface, and these take their turn for a while at putting forth the characteristic cyathia and cymes, on peduncles which are variably very short or almost absurdly long. Under cultivation the branches are rarely drawn underground, but they pile up in a fantastic medley, each branch differing somewhat from the others in shape, globose, clavate or cylindric, so that a large potted plant looks not unlike a bowl of quaint, irregular marbles or tangled strings of misshapen large beads. Occasionally the longer peduncles produce little new branches up aloft, instead of the expected inflorescence, until of their own weight these adventitious newcomers drop over to the ground, send out rootlets and begin life on their own accord.”
Inflorescence
Here I would first like to discuss the special inflorescence of Euphorbias. What we see as a flower is in fact a whole bouquet. Each stamen is a separate male flower stripped of all non-essential parts. Only the stamen itself, ending in 2 anthers, is therefore left. In the same way, the pistil is in itself a separate female flower. Pistil and stamens grow from a kind of cup, called involucrum. On the outside of the involucrum there are usually 4 or 5 nectar glands. All those basic flowers, together with what is around them, form what an ordinary plant enthusiast sees as ‘the flower’. But in botany, this combination of flowers is called a cyathium.
Fig. 3
In figure 3 we see a young stage of the very special inflorescence of E. globosa. In the middle you can see the pistil with the 3 stigma lobes and around it the anthers in an immature stage. The diameter of this inflorescence at that time was 10 mm. If you look at that picture you will immediately understand why Haworth chose the name ‘Dactylanthes’ (finger-flower) for this type of plant.
The ‘fingers’ are outgrowths of the nectar glands. The nectar is secreted by the wart-like elevations. In E. globosa we usually find 3 or 4, sometimes 5 ‘fingers’ per nectar gland.
Fig. 4
In figure 4 we see an inflorescence at a much later stage. The pistil has reached its final size and the stigma with its 3 lobes sticks out between 2 fingers. The swollen ovary is clearly visible. At the bottom left of the cup we see 2 immature anthers and at the top right a stamen with 2 ripe, pollen producing anthers. If you look closely, you can see about halfway up this stamen, at the level of the cup edge, a kind of ring around the stamen. That is the transition from flower stalk to stamen. When the stamen (= flower) has finished flowering, it drops off its flower stalk. What remains is a kind of hollow tube. In the cup we also see a number of flower stalks from which the finished stamen has already broken off. On the 3 fingers of the honey gland at the bottom right, we see a few of those broken stamens.
A peculiar thing in the photo is that at the upper nectar gland there were first 3 fingers of which the middle one has divided again halfway.
It is reported that at the ends of the flowering stems sometimes not cyathia, but small spherical sections develop. If the stems bend due to the weight of the segments and touch the soil, they will quickly take root.
In cultivation I bought my oldest E. globosa around 1990 at a cactus market. It was probably already a slightly older plant, consisting of some larger gray-white balls, and I wondered if it had been imported. This plant has now grown into a collection of perhaps 70 stems, covered with a tangle of dried flowering branches which makes it impossible to make a nice photo of it. This plant is self-fertile. Usually, after flowering, the fruits with 3 fertile seeds -characteristic for Euphorbias- appear .
Fig. 5, a seedling of a few years old
In figure 6, we see seedlings of 5 months old in a pot.
In figure 7, photographed when the same seedlings were 4 months old, it can be seen that one of the plants in the same pot is already preparing to flower.
Euphorbia globosa requires a sunny place with not too much water. If not, the spheres lengthen, and the plant will look like the closely related E. ornithopus.(The same group also includes E. tridentata, E. polycephala, E. wilmaniae and E. planiceps).
Minimum temperature in winter 8 ºC, although in literature it is reported that the plant can withstand frost.
Finally In a series of articles about the plants grown in the botanical garden in Geneva, Alphonse de Candolle wrote an extensive article about E. globosa in 1836. There was a beautiful folding plate in which all parts of the plant were depicted in detail. See below. Unfortunately, the scan is slightly distorted on the left-hand side.
Literature Buddensiek, V. (1998). Sukkulente Euphorbien; 25, 101.
Candolle, A. P. de. (1836). Septième Notice sur les Plantes rares cultivées dans le Jardin de Genève, Memoires de la Société de physique et d’histoire naturelle de Genève 7 (2): 288 – 293, fig. 5.
Haworth, A. H. (1812). Synopsis plantarum succulentarum: 132.
Haworth, A. H. (1823). Plantae rarae Succulentae; a Description of some rare Succulent Plants, The Philosophical Magazine and Journal 62: 382.
Sims, J. (1926). Euphorbia globosa, Curtis’s Botanical Magazine 53, plaat 2624.
White, A; Dyer, R.A. and Sloane, B.L. (1941). The succulent Euphorbieae (Southern Africa).
This species was discovered in 1960 on the lower slopes (700–1200 m.) of Mt. Kasigau in southern Kenya, where it grows on exposed rock faces. It has not been recorded from other places since then.
Although at first sight it reminds one of a multi-angled E. heterochroma, it has some distinguishing features: usually not higher than 1 m, branches always rigid and uniform sap-green.
The plants do very well in cultivation and can easily be propagated from cuttings.
It is a shrub with more or less erect branches which are sparsely rebranched, prominently 6–9-angled, 1–3 cm. thick and bright green; the angles are shallowly toothed, with teeth 6–12 mm. apart.
The grey spines are to 8 mm. long; without or with rudimentary prickles
Cyathia are ± 3 × 4 mm., with cup-shaped involucres; the golden yellow glands are transversely oblong, ± 1 × 2 mm. and the lobes are subcircular, ± 1.3 × 1.3 mm.
The capsules are obtusely 3-lobed and exserted on a reflexed pedicel to 4 mm. long.
Many Crassulas are beautiful mimicry plants. This one owes its name deceptor (= impostor) to the fact that the gray-green rosettes set with small dots look like small angular stones and in their habitat in southern Namibia and Namaqualand are hidden amongst the quartz stones that occur there.
The species was found in 1897 by Alston (of Avonia alstonii fame) and described by Schönland & Baker in 1902. Schönland later changed the name to Crassula deceptrix. This suggests that he considered cheating to be a trait that suits women more than men. But the real reason was that the word Crassula is feminine, and he thought (wrongly) that the species name had to be feminine as well. Later (1974) the name change was revoked.
Crassula deceptor is variable in shape and size. In nature, the rosettes are about 2.5 cm in diameter and up to about 8 cm tall. The plant in figure 1 has the same width.
Fig. 1: Crassula deceptor resembles a jagged piece of stone
The stems divide dichotomously and form a compact cluster over time.
For me in the Netherlands, the plant always blooms around October. As is the case with many Crassulas, this is not a spectacular but nevertheless graceful sight. The plants form branched flowering stems which protrude well above them and produce several tiny cream-green flowers which turn brown after flowering (fig. 2).
Fig. 2: Crassula deceptor in bloom
As for its cultivation: full sun, little water in summer and none in winter. With a less sunny location and/or a lot of water, they do not retain the compact shape. Be careful with water staying behind on the rosettes.
C. cornuta (figure 3), also described by Schönland, is nowadays considered to be synonymous with C. deceptor. Yet it clearly deviates from the standard form of the species. The leaves are longer, more pointed and much lighter in colour.
Fig 3.
For the sake of completeness, it is worth mentioning that there are also quite a few hybrids in which C. deceptor is one of the parents. I have a hybrid of C. deceptor x C. susannae (fig. 4); the diameter of this plant is just 2 cm.
Fig 4
In ‘Crassula’ by Gordon Rowley, this cross is called ‘Dorothy’. He also mentions the following hybrids: ‘Frosty’ (C. deceptor x tecta); ‘Gandalf’ (C.deceptor x mesembryanthemopsis); ‘Moonglow’ (C. deceptor x perfoliata var. falcata); ‘Shogun’ (C. deceptor x hemisphaerica) and the multihybrid ‘Star Child’ (C. deceptor x ‘Starbust’), ‘Starbust’ being a hybrid of C. ausensis x pyramidalis.
Fig. 5: Leaves of C. ‘Frosty’
Fig.6: Flowers of C. ‘Frosty’
Fig. 7: C. deceptor in habitat
Literature: B.K. Boom (1980), De Crassula’s van onze collecties, Succulenta 59 [8]: 176-179 Mia C. Karsten (1941), Zuid-Afrikaansche succulente reisherinneringen I, de botanische tuin te Stellenbosch (6), Succulenta 23 [6]: 65
Gordon Rowley (2003), Crassula, Cactus & Co
First published in Succulenta 89 [2]: febr. 2010. Translation from Dutch: FN.
“The most magnificent and beautiful of all the Crassulaceae is the scarlet Rochea coccinea. Emerging from crevices or growing freely on the flat rocks, the large perennials accommodate their root system in a layer of soil only a few centimeters thick, which sits so loosely on the rock that it can be lifted off together with the plant. The fact that despite its fleshy leaves it is dependent on the moisture coming from the summer clouds is best demonstrated by the cultivation experiment in Cape Town: it only thrives there, even moved to similar locations, if it is watered at least once in a while in summer.” This is a quote from the book ‘Das Kapland’ by Rudolf Marloth, published in 1908.
According to literature, C. coccinea grows at altitudes over 800 meters in the Southwest Cape. As distribution area, the Cape Peninsula (especially Table Mountain near Cape Town) and the area between Paarl and Bredasdorp (135 km to the southeast) are mentioned.
In reality the area is considerably larger, as evidenced by the photo taken by Bertus Spee in mid-November 2008 in Citrusdal, 125 km north of Paarl. This was at the beginning of the flowering period. In nature it is part of the so-called Fynbos vegetation.
Fig. 1 C. coccinea and an Adromischus in 2008 at the beginning of the flowering period (mid-November) in Citrusdal (Western Cape). Photo Bertus Spee.
The stems become about 40 cm high and they branch from the base. In older plants, the stems become bare and brown from below, with only the tips bearing leaves. Flowering occurs in summer (in Europe in winter). Due to the long flower tube of about 4 cm, only butterflies can access the nectar. Back in 1925 Marloth noted that the butterfly Meneris (now Aeropetes) tulbaghia is the main pollinator. This butterfly is called “the table mountain beauty” or “the mountain pride butterfly” in South Africa and is specialized in red flowers. It is rumoured that you will be chased by the butterfly if you wear a red cap. It is probably the only pollinator for at least 15 plant species in fynbos. Among those 15 are 3 species of the genus Cyrtanthus, which is also known to succulent lovers, and 7 of Gladiolus.
History Like Kumara (Aloe) plicatilis , discussed in an earlier post, Crassula coccinea belongs to the succulents that were in cultivation in the Netherlands as early as the late seventeenth century. It is first mentioned in a book from 1706 by Caspar Commelin about rare plants in the Hortus Medicus, the predecessor of the current Hortus Botanicus in Amsterdam. The ‘name’ that Commelin gave was ‘Cotyledon Africana frutescens flore umbellato Coccineo’ which means something like: bushy growing African Cotyledon with scarlet flowers in a flat-topped inflorescence.
The accompanying black-and-white drawing shows a plant with flower buds about to open.
Fig. 2 Image of Crassula coccinea in Casper Commelin’s 1706 book.
In the Moninckx atlas (see also the earlier post about Kumara plicatilis) there is also a beautiful colour image. According to the University of Amsterdam, this was made somewhere between 1686 and 1706 by Maria Moninckx. Although the images clearly differ from each other, they both seem to show the same specimen.
Fig . 3 Watercolour of Crassula coccinea by Maria Moninckx (made between 1686 and 1706). The image in the book ‘Phytanthoza iconographia’ by Johann Wilhelm Weinman from 1737 is also beautiful. Here C. coccinea is depicted together with C. perfoliata and C. tetragona. The funny thing is that a mistake was made with the Latin specific name. It says “fructescens” (bearing fruit) instead of “frutescens” (shrub-forming). Not just a clerical error, because in the German text the plant is called “Fruchtbringend”.
Fig. 4 Image of Crassula coccinea (a), C. perfoliata (b) and C. tetragona (c), in Phytanthoza iconographia from 1739.
Nomenclature
As far as I can tell, the name Crassula first appeared in 1732. The German botanist Johann Jakob Dillenius used it for eight succulents in his work ‘Hortus Elthamensis’. The name means something like ‘fat one’. The common name is ‘thick leaf’. Linnaeus adopted the genus name in his ‘Genera Plantarum’ in 1735 and in 1753 he already described 10 species of Crassula in his ‘Species Plantarum’ (first edition). It’s curious that he mentions Ethiopia as the place of origin for Crassula coccinea, as well as for 7 other Crassulas. Of the 2 remaining ones, C. alternifolia is only given a question mark and in the case of C. scabra, he mentions “Habitat in Africa?”.
Later on, C. coccinea was placed in the genus Rochea. A.P. de Candolle created this genus in 1802 to accommodate the Crassulas with large tubular flowers. The genus name refers to the Swiss botanist Daniel de la Roche. In 1805 the South African mycologist Christiaan Hendrik Persoon changed the genus name to Larochea, after which the Austrian Leopold Trattinnick (botanist and also mycologist) changed it into Dietrichia in 1812. Adrian Haworth in his turn created the genus Kalosanthes in 1821 to accommodate 8 Crassulas including C. coccinea.
He also reported a white-flowered variety (“floribus albis”) based on a book with drawings and descriptions of succulent plants by Richard Bradley from 1716. Unfortunately, a number of drawings in that book were incorrectly numbered and that resulted in, among other things, C. coccinea containing the text of C. tetragona. And there was talk of “small white flowers”. One wonders how well Haworth looked at his sources, as C. coccinea has the largest flowers of the entire Crassula genus.
All the above genus names have been declared invalid and it is again just Crassula. Kalosanthes has remained as the name for one of the 20 sections within the genus Crassula. This section contains only 3 of the over 200 Crassula species. Apart from C. coccinea they are C. fascicularis (= C. odoratissima) and C. obtusa (= C. jasminea). See also fig. 5, where the species of the section are depicted.
Fig. 5 In addition to Tylecodon reticulatus, the 3 Crassulas from the current Kalosanthes section are also pictured. Way back in 1912 Marloth indicated that C. coccinea for its pollination depends on the butterfly Menera (Aeropetes) tulbaghia.
Characteristic of the plants in this section are the long tubular flowers which flare out saucer-shaped at the end. This shape is caused by the fact that the 5 petals are fused over at least half their length and have a widened end at right angles.
In South Africa, C. coccinea is called ‘klipblom’ (stone flower). Also common is the name “Red Crassula” and in older literature one may come across the name ’emperor’s crown’.
Cultivation C. coccinea was once a widely cultivated indoor plant. In Victorian times, the plant was loved in England for its large bright red flowers and it is one of the few succulents that has been grown commercially on a large scale . The other 2 crassulas from the Kalosanthes section were also widely grown in England at that time. C. obtusa under the name Rochea jasminea -referring to the jasmine-like white flowers and C. fascicularis under the name Rochea odoratissima because of the strong scent.
In Germany too people were involved in growing and breeding this species. Gordon Rowley in his book ‘Crassula, A Grower’s Guide’ mentions that the famous Haage firm in Erfurt offered three cultivars in their 1857 catalogue. In 1868 there were already ten. They have all disappeared from the collections again.
Among the succulent plant lovers we don’t see C. coccinea very much anymore. Young plants are still fresh and compact, but soon they become too tall and untidy. Nevertheless, the species is still grown professionally.
It grows easily and quickly in well-drained soil. Often a mixture of two parts of leaf mould, one part clay and one part sand is recommended. In full sun, a lot of water and fertilizer is required. The plants can be grown from seed or cuttings. Seedlings can already bloom in the first year. The plants are susceptible to the fungal disease ‘rust’ but can’t stand fungicides (pesticides against fungi). After a few years, the plants become ugly with dying leaves at the bottom and deflecting shoots. That is the time to take cuttings. In a mixture of peat and sand they will quickly root.
Cuttings can also just be put in a jar of water. Once they are growing, heading them several times is recommended. This creates compact bushy plants. Supplying a larger pot may also be needed regularly. In summer they can stay outside without problems. In winter keep them cool but do not let them dry out.
Fig. 6 Crassula coccinea flowering in cultivation. The species was massively grown in the 18th century for its large, bright red flowers.
Hybrids In comparison to some other Crassulas, C. coccinea has not often been used for hybridisation. Probably the oldest hybrid is C. coccinea x falcata. This is midway in appearance between the two species and was named Kalorochea langleyensis. In1898, a specimen was exhibited at the Royal Horticultural Society’s show in Chelsea. This hybrid, which according to the current rules should be called Crassula x langleyensis does not seem to be cultivated anymore.
A hybrid with C. undulata was also once produced, but has suffered the same fate and the same applies to the cross with the white-flowering C. obtusa (Rochea jasminea).
Literature Arianoutsou-Faraggitaki, M & Groves, R, H. (2012). Plant-animal interactions in Mediterranean-type ecosystems: 137 – 147.
Candolle, A.P. de & Redouté, P.J. (1799-1837): Plantarum Historia Succulentarum, 1: 1.
Commelin, C. (1706). Horti medici amstelædamensis plantæ rariores et exoticæ ad vivum æri incisæ: 24.
Dillenius, J. (1732): Hortus Elthamensis 1: 114 – 121.
Knippels, P.J.M. (1992). The household consumption and turnover of succulents at the flower auctions, Succulenta 71 (1): 33 -38.
Linnaeus, C. (1737). Genera Plantarum: 89.
Linnaeus, C. (1753). Species Plantarum 1: 282 -283.
Marloth, R. (1908). Das Kapland, Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer “VaIdivia” 1898-1899, bd.2, T3:142, 339.
Marloth, R. (1925). The Flora of South-Africa 2: 17 – 24.
Rowley, G. ( 2003): Crassula, A Grower’s Guide.
Weinmann, J.W. (1739) Phytanthoza iconographia 2: 435.
Originally published in Succulenta June 2016. Translated from Dutch by FN.












 Fig 1
Fig 1
 Fig 4 C. columnaris ssp. prolifera in the Knersvlakte (photo by Sofia Etzold, with thanks to “Photo Guide to Plants of Southern Africa”). In Afrikaans, this variety is called ‘sentkannetjie’. ‘Sent’ in this expression is derived from the English ‘scent’ and kannetjie stands for bottle. This does justice to both the plant’s shape and the flowers’ sweet smell.
Fig 4 C. columnaris ssp. prolifera in the Knersvlakte (photo by Sofia Etzold, with thanks to “Photo Guide to Plants of Southern Africa”). In Afrikaans, this variety is called ‘sentkannetjie’. ‘Sent’ in this expression is derived from the English ‘scent’ and kannetjie stands for bottle. This does justice to both the plant’s shape and the flowers’ sweet smell.






































 Fig. 2: Crassula deceptor in bloom
Fig. 2: Crassula deceptor in bloom Fig 3.
Fig 3.